2qjm: Difference between revisions
m Protected "2qjm" [edit=sysop:move=sysop] |
No edit summary |
||
| (6 intermediate revisions by the same user not shown) | |||
| Line 1: | Line 1: | ||
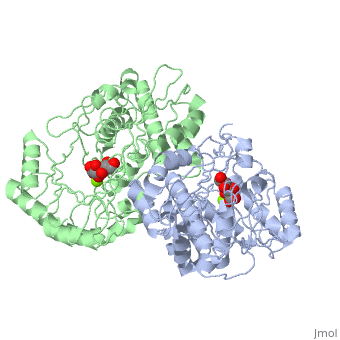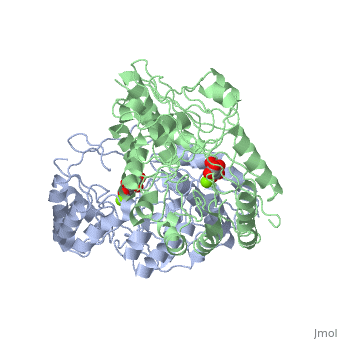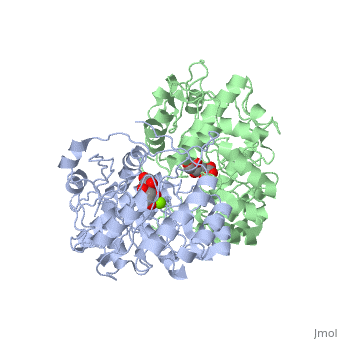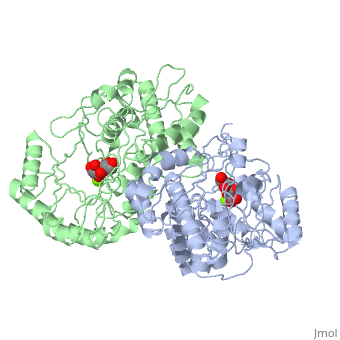
==Crystal structure of the K271E mutant of Mannonate dehydratase from Novosphingobium aromaticivorans complexed with Mg and D-mannonate== | |||
<StructureSection load='2qjm' size='340' side='right'caption='[[2qjm]], [[Resolution|resolution]] 2.20Å' scene=''> | |||
== Structural highlights == | |||
<table><tr><td colspan='2'>[[2qjm]] is a 4 chain structure with sequence from [https://en.wikipedia.org/wiki/Novosphingobium_aromaticivorans Novosphingobium aromaticivorans]. Full crystallographic information is available from [http://oca.weizmann.ac.il/oca-bin/ocashort?id=2QJM OCA]. For a <b>guided tour on the structure components</b> use [https://proteopedia.org/fgij/fg.htm?mol=2QJM FirstGlance]. <br> | |||
</td></tr><tr id='method'><td class="sblockLbl"><b>[[Empirical_models|Method:]]</b></td><td class="sblockDat" id="methodDat">X-ray diffraction, [[Resolution|Resolution]] 2.2Å</td></tr> | |||
<tr id='ligand'><td class="sblockLbl"><b>[[Ligand|Ligands:]]</b></td><td class="sblockDat" id="ligandDat"><scene name='pdbligand=CS2:D-MANNONIC+ACID'>CS2</scene>, <scene name='pdbligand=MG:MAGNESIUM+ION'>MG</scene></td></tr> | |||
<tr id='resources'><td class="sblockLbl"><b>Resources:</b></td><td class="sblockDat"><span class='plainlinks'>[https://proteopedia.org/fgij/fg.htm?mol=2qjm FirstGlance], [http://oca.weizmann.ac.il/oca-bin/ocaids?id=2qjm OCA], [https://pdbe.org/2qjm PDBe], [https://www.rcsb.org/pdb/explore.do?structureId=2qjm RCSB], [https://www.ebi.ac.uk/pdbsum/2qjm PDBsum], [https://prosat.h-its.org/prosat/prosatexe?pdbcode=2qjm ProSAT]</span></td></tr> | |||
</table> | |||
== Function == | |||
[https://www.uniprot.org/uniprot/MAND_NOVAD MAND_NOVAD] Catalyzes the dehydration of D-mannonate. Has no detectable activity with a panel of 70 other acid sugars (in vitro).<ref>PMID:17944491</ref> <ref>PMID:24697546</ref> | |||
== Evolutionary Conservation == | |||
[[Image:Consurf_key_small.gif|200px|right]] | |||
Check<jmol> | |||
<jmolCheckbox> | |||
<scriptWhenChecked>; select protein; define ~consurf_to_do selected; consurf_initial_scene = true; script "/wiki/ConSurf/qj/2qjm_consurf.spt"</scriptWhenChecked> | |||
<scriptWhenUnchecked>script /wiki/extensions/Proteopedia/spt/initialview01.spt</scriptWhenUnchecked> | |||
<text>to colour the structure by Evolutionary Conservation</text> | |||
</jmolCheckbox> | |||
</jmol>, as determined by [http://consurfdb.tau.ac.il/ ConSurfDB]. You may read the [[Conservation%2C_Evolutionary|explanation]] of the method and the full data available from [http://bental.tau.ac.il/new_ConSurfDB/main_output.php?pdb_ID=2qjm ConSurf]. | |||
<div style="clear:both"></div> | |||
<div style="background-color:#fffaf0;"> | |||
== Publication Abstract from PubMed == | |||
The d-mannonate dehydratase (ManD) function was assigned to a group of orthologous proteins in the mechanistically diverse enolase superfamily by screening a library of acid sugars. Structures of the wild type ManD from Novosphingobium aromaticivorans were determined at pH 7.5 in the presence of Mg2+ and also in the presence of Mg2+ and the 2-keto-3-keto-d-gluconate dehydration product; the structure of the catalytically active K271E mutant was determined at pH 5.5 in the presence of the d-mannonate substrate. As previously observed in the structures of other members of the enolase superfamily, ManD contains two domains, an N-terminal alpha+beta capping domain and a (beta/alpha)7beta-barrel domain. The barrel domain contains the ligands for the essential Mg2+, Asp 210, Glu 236, and Glu 262, at the ends of the third, fourth, and fifth beta-strands of the barrel domain, respectively. However, the barrel domain lacks both the Lys acid/base catalyst at the end of the second beta-strand and the His-Asp dyad acid/base catalyst at the ends of the seventh and sixth beta-strands, respectively, that are found in many members of the superfamily. Instead, a hydrogen-bonded dyad of Tyr 159 in a loop following the second beta-strand and Arg 147 at the end of the second beta-strand are positioned to initiate the reaction by abstraction of the 2-proton. Both Tyr 159 and His 212, at the end of the third beta-strand, are positioned to facilitate both syn-dehydration and ketonization of the resulting enol intermediate to yield the 2-keto-3-keto-d-gluconate product with the observed retention of configuration. The identities and locations of these acid/base catalysts as well as of cationic amino acid residues that stabilize the enolate anion intermediate define a new structural strategy for catalysis (subgroup) in the mechanistically diverse enolase superfamily. With these differences, we provide additional evidence that the ligands for the essential Mg2+ are the only conserved residues in the enolase superfamily, establishing the primary functional importance of the Mg2+-assisted strategy for stabilizing the enolate anion intermediate. | |||
Evolution of enzymatic activities in the enolase superfamily: D-Mannonate dehydratase from Novosphingobium aromaticivorans.,Rakus JF, Fedorov AA, Fedorov EV, Glasner ME, Vick JE, Babbitt PC, Almo SC, Gerlt JA Biochemistry. 2007 Nov 13;46(45):12896-908. Epub 2007 Oct 18. PMID:17944491<ref>PMID:17944491</ref> | |||
From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine.<br> | |||
</div> | |||
<div class="pdbe-citations 2qjm" style="background-color:#fffaf0;"></div> | |||
== | ==See Also== | ||
[[ | *[[Mandelate racemase/muconate lactonizing enzyme|Mandelate racemase/muconate lactonizing enzyme]] | ||
*[[Mandelate racemase/muconate lactonizing enzyme 3D structures|Mandelate racemase/muconate lactonizing enzyme 3D structures]] | |||
== | == References == | ||
< | <references/> | ||
__TOC__ | |||
</StructureSection> | |||
[[Category: Large Structures]] | |||
[[Category: Novosphingobium aromaticivorans]] | [[Category: Novosphingobium aromaticivorans]] | ||
[[Category: Almo | [[Category: Almo SC]] | ||
[[Category: Fedorov | [[Category: Fedorov AA]] | ||
[[Category: Fedorov | [[Category: Fedorov EV]] | ||
[[Category: Gerlt | [[Category: Gerlt JA]] | ||
[[Category: Rakus | [[Category: Rakus JF]] | ||
[[Category: Vick | [[Category: Vick JE]] | ||
Latest revision as of 14:34, 30 August 2023
Crystal structure of the K271E mutant of Mannonate dehydratase from Novosphingobium aromaticivorans complexed with Mg and D-mannonateCrystal structure of the K271E mutant of Mannonate dehydratase from Novosphingobium aromaticivorans complexed with Mg and D-mannonate
Structural highlights
FunctionMAND_NOVAD Catalyzes the dehydration of D-mannonate. Has no detectable activity with a panel of 70 other acid sugars (in vitro).[1] [2] Evolutionary Conservation Check, as determined by ConSurfDB. You may read the explanation of the method and the full data available from ConSurf. Publication Abstract from PubMedThe d-mannonate dehydratase (ManD) function was assigned to a group of orthologous proteins in the mechanistically diverse enolase superfamily by screening a library of acid sugars. Structures of the wild type ManD from Novosphingobium aromaticivorans were determined at pH 7.5 in the presence of Mg2+ and also in the presence of Mg2+ and the 2-keto-3-keto-d-gluconate dehydration product; the structure of the catalytically active K271E mutant was determined at pH 5.5 in the presence of the d-mannonate substrate. As previously observed in the structures of other members of the enolase superfamily, ManD contains two domains, an N-terminal alpha+beta capping domain and a (beta/alpha)7beta-barrel domain. The barrel domain contains the ligands for the essential Mg2+, Asp 210, Glu 236, and Glu 262, at the ends of the third, fourth, and fifth beta-strands of the barrel domain, respectively. However, the barrel domain lacks both the Lys acid/base catalyst at the end of the second beta-strand and the His-Asp dyad acid/base catalyst at the ends of the seventh and sixth beta-strands, respectively, that are found in many members of the superfamily. Instead, a hydrogen-bonded dyad of Tyr 159 in a loop following the second beta-strand and Arg 147 at the end of the second beta-strand are positioned to initiate the reaction by abstraction of the 2-proton. Both Tyr 159 and His 212, at the end of the third beta-strand, are positioned to facilitate both syn-dehydration and ketonization of the resulting enol intermediate to yield the 2-keto-3-keto-d-gluconate product with the observed retention of configuration. The identities and locations of these acid/base catalysts as well as of cationic amino acid residues that stabilize the enolate anion intermediate define a new structural strategy for catalysis (subgroup) in the mechanistically diverse enolase superfamily. With these differences, we provide additional evidence that the ligands for the essential Mg2+ are the only conserved residues in the enolase superfamily, establishing the primary functional importance of the Mg2+-assisted strategy for stabilizing the enolate anion intermediate. Evolution of enzymatic activities in the enolase superfamily: D-Mannonate dehydratase from Novosphingobium aromaticivorans.,Rakus JF, Fedorov AA, Fedorov EV, Glasner ME, Vick JE, Babbitt PC, Almo SC, Gerlt JA Biochemistry. 2007 Nov 13;46(45):12896-908. Epub 2007 Oct 18. PMID:17944491[3] From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine. See Also
References
|
| ||||||||||||||||||