1l6j
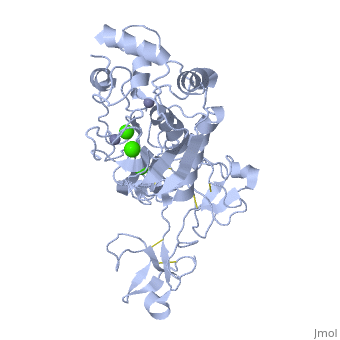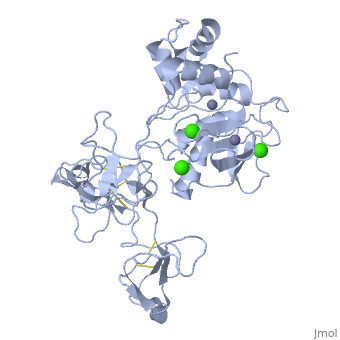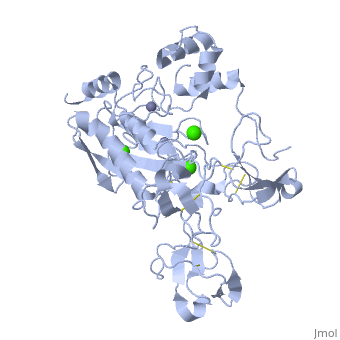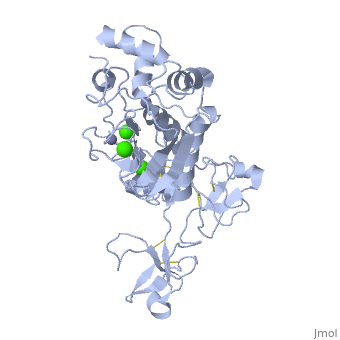
Crystal structure of human matrix metalloproteinase MMP9 (gelatinase B).Crystal structure of human matrix metalloproteinase MMP9 (gelatinase B).
Structural highlights
DiseaseMMP9_HUMAN Defects in MMP9 may be a cause of susceptibility to intervertebral disc disease (IDD) [MIM:603932; also known as lumbar disk herniation (LDH). IDD is one of the most common musculo-skeletal disorders and the predominant cause of low-back pain and unilateral leg pain.[1] Defects in MMP9 are the cause of metaphyseal anadysplasia type 2 (MANDP2) [MIM:613073. Metaphyseal anadysplasia consists of an abnormal bone development characterized by severe skeletal changes that, in contrast with the progressive course of most other skeletal dysplasias, resolve spontaneously with age. Clinical characteristics are evident from the first months of life and include slight shortness of stature and a mild varus deformity of the legs. Patients attain a normal stature in adolescence and show improvement or complete resolution of varus deformity of the legs and rhizomelic micromelia. FunctionMMP9_HUMAN May play an essential role in local proteolysis of the extracellular matrix and in leukocyte migration. Could play a role in bone osteoclastic resorption. Cleaves KiSS1 at a Gly-|-Leu bond. Cleaves type IV and type V collagen into large C-terminal three quarter fragments and shorter N-terminal one quarter fragments. Degrades fibronectin but not laminin or Pz-peptide.[2] Evolutionary Conservation Check, as determined by ConSurfDB. You may read the explanation of the method and the full data available from ConSurf. Publication Abstract from PubMedThe X-ray crystal structure of the proform of human matrix metalloproteinase MMP9 has been solved to 2.5 A resolution. The construct includes the prodomain, the catalytic domain and three FnII (fibronectin type II) domains. The prodomain is inserted into the active-site cleft, blocking access to the catalytic zinc. Comparison with the crystal structure of the most closely related MMP, MMP2, indicates that the conformations of residues in the active-site cleft and in the cysteine-switch peptide of the prodomain are highly conserved and that design of MMP9-specific inhibitors will be challenging. In common with MMP2, the MMP9 S1' inhibitor-binding pocket is large compared with that of other MMPs. One small point of difference in the S1' binding pockets of MMP9 and MMP2 may provide an opportunity to explore the design of specific inhibitors. The side chain of Arg424 in MMP9 is angled slightly away from the S1' pocket when compared with the corresponding residue in MMP2, Thr424. The secondary structure of the FnII domains is conserved between the two closely related MMPs, although the second FnII domain makes no contact with the catalytic domain in MMP9, while the same domain in MMP2 has a substantial area of interaction with the catalytic domain. Structure of the C-terminally truncated human ProMMP9, a gelatin-binding matrix metalloproteinase.,Elkins PA, Ho YS, Smith WW, Janson CA, D'Alessio KJ, McQueney MS, Cummings MD, Romanic AM Acta Crystallogr D Biol Crystallogr. 2002 Jul;58(Pt 7):1182-92. Epub 2002, Jun 20. PMID:12077439[3] From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine. See AlsoReferences
|
| ||||||||||||||||||