Replication termination in E. coli and B. subtilis
ter elements, Tus and RTPter elements, Tus and RTP
Bacterial DNA is circular and is replicated by two self-sufficient replication forks, progressing in opposite directions. These forks contain several factors, such as a helicase and single stranded-DNA binding proteins, involved in unwinding and maintaining the separation of the 2 parent strands whilst daughter strands are synthesised. The life cycle of bacteria depends upon the coordinated termination of this DNA replication [11].
ter (terminator) elements are asymmetric patterns of DNA that act as protein binding sites [1]. These elements are situated in the terminus region, approximately opposite the origin of replication [11]. The binding of specific proteins to ter elements provides a "trap" for the proceeding replication fork, catching the replication fork as it passes. There are several ter elements responsible for stopping each replication fork, with each of these elements being specific for the fork passing in one direction only, that is, they have functional polarity [3,11]. The ter element responsible for catching the clockwise replication fork will allow the anticlockwise fork to proceed unchecked, until it is stopped by its own anticlockwise facing ter element fork trap [2,3].
Replication fork traps have been identified in multiple species possessing circular chromosomes, including Escherichia coli, Bacillus subtilis and more recently Salmonella typhimurium [2,9]. Fork traps prevent over replication of the bacterial chromosome and stall a faster fork in the case that one side of the replication was proceeding faster than the other. The presence of several ter sites for each replication fork is necessary to ensure that replication termination occurs and indicates a sense of redundancy, supported by the highly conserved nature of ter sites and their highly specific cognate binding capabilities.
E. coli and TusE. coli and Tus
|
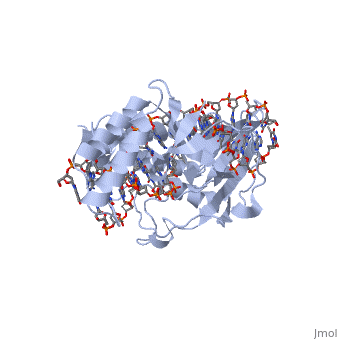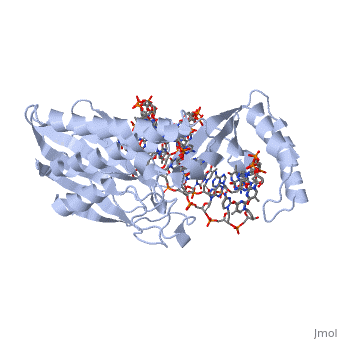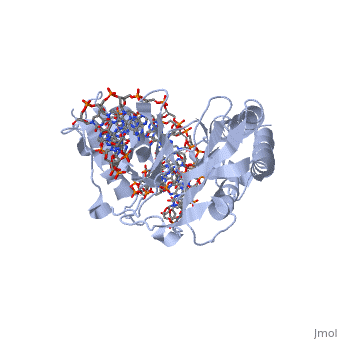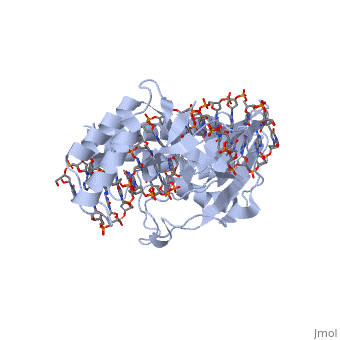
E. coli posess two clusters of five ter elements, named ter A-J, each of which is 23 base pairs long [3]. They are situated around 100kB either side of the termination region. The cognate binding protein to E. coli ter elements is the Tus (Termination utilisation substance) protein.
Tus is a . It acts by binding to the ter site and inhibiting the helicase coupled strand separation function of DnaB helicase in the replication machinery [6]. This prevents progression of the DNA replication machinery through the ter site and contributes to replicative arrest. The TUS protein is composed of . This structure forms a positively charged central cleft that can accomodate 13 base pairs of duplex DNA. Tus binds and acts monomerically, and has exhibited strong binding affinity [3].
B. subtilis and RTPB. subtilis and RTP
|
B.subtilis posesses nine ter elements of 29 base pair sequences containing 16 base pair imperfectly inverted repeats [3]. These are highly conserved sequences, which create functional A and B sites. The B site represents a more strongly acting binding core site, whilst the A site represents an auxiliary binding domain with lesser affinity for the RTP molecule.
The RTP protein is a 29 kDa member of the winged helix family and consists of . RTP acts as a . Two RTP monomers join to form a dimer which binds to either the A or B site of a ter element. Even though the protein and its binding at each site is the same, the stronger B site is always filled before the A, and both sites must be filled for fork arrest to occur [4,5].
As in E. coli, the B. subtilis fork arrest process is mono-directionally selective. If the replication fork arrives at the A site before the B site, it will pass through unhindered, however if the B site is reached before the A site, the fork is arrested and termination ensues [5,8].
The effect of knockout of ter elements or their cognate binding proteins; the real function of terThe effect of knockout of ter elements or their cognate binding proteins; the real function of ter
It has been found that the ter site DNA in E. coli and B. subtilis does not bear any sequence homology [11]. Similarly, the termination proteins Tus and RTP do not share conformational characteristics, three dimensional homology or binding similarities [12]. The only similarity identifyable is their shared biological function, which appears to be an adaption to long term survival with a circular chromosome [4]. Several early experiments showed that under laboratory conditions, the knockout of either rtp or tus genes (leading to loss of protein synthesis) does not cause an observable phenotype in either B. subtilis or E. coli [8,9].
The presence of the fork trap constructs has several important and advantageous consequences for the organism in question. These include:
- Due to the high conservation of sequences within a species, the presence of multiple trap regions introduces a level of redundancy, whereby if a single base mutation in the ter element was to inactivate the region, another ter element further towards the terminus-to-origin direction might be used. [1]
- Multiple ter sites allow for a level of speed regulation, such that the faster of 2 replication forks might be slowed down when progressing faster than the other. This might occur if one side of the replicating chromosome had to pause to allow DNA repair mechanisms to be completed. [10]
However these advantages do not explain the developmental pressures leading to the development of these systems individually, nor do they explain why the removal of activity of these sites by knockout causes no functional phenotype.
The functional significance of the replication fork trap construct is that without it, replication would not be forced to terminate at 180˚ from the origin, and it may continue back in the terminus-to-origin direction. The development of a fork trap construct in circular chromosomes suggests that this would be undesirable for the organism. Reasons for this may include the fact that the majority of transcribed and translated genes are oriented for transcription in origin-to-terminus direction. If replication machinery was allowed to continue on in a terminus-to-origin orientation, there would be the potential for head-on-collision between transcription and replication machinery, which has been proven in the past to have deleterious affects [4].
More recent studies have showed a highly important and genome wide regulatory role for the ter sites and their cognate binding proteins. Study of E. coli shows that when mutations or knockouts are introduced to DNA polymerase A, the loss of function of the ter sites leads to increased levels of DNA overproduction. Furthermore, cells with Tus-terB deletions also exhibited increased rates of DNA overproduction. When Tus protein was provided to such cells, this overproduction was corrected, confirming that the absence of Tus (and not the loss of polA function) was responsible for the DNA overproduction [6]. Similar studies in B. subtilis show that when mutations are introduced to partitioning genes in combination with mutation to the rtp gene, an increase in anucleate cell production results. Partitioning genes are genes responsible for the accurate separation of replication products into daughter cells, and include the proteins spoIIIE and ripX. B. subtilis studies show that whilst the loss of rtp does not cause partitioning defects in wild-type background, when combined with partitioning defects an increase in anucleate cell production results [5]. These studies suggest a more global role for the the ter sites and their cognate binding proteins, and suggests their global responsibility for maintainance of the termination of replication as a safeguard against the affects of mutations in the highly important replication machinery.
ReferencesReferences
1. Duggin, I and Bell, S (2009) Termination structures in the Escherichia coli chromosome replication fork trap. J. Mol. Biol. 387:532-539.
2. Kamada et al. (1996) Structure of a replication-terminator protein complexed with DNA. Nature 387.
3. Vivian et al. (2007) An asymmetric structure of the Bacillus subtilis replication terminator protein in complex with DNA. J. Mol. Biol. 370:481-491.
4. Duggin et al. (2008) The replication fork trap and termination of chromosome replication. J. Mol. Biol.. 70(6):1323-1333.
5. Lemon et al. (2000) Effects of replication termination mutants on chromosome partitioning in Bacillus subtilis. PNAS 98(1):212-217.
6. Markovitz, A. (2005) A new in vivo termination function for DNA polymerase I of Escherichia coli K12. Molecular Microbiology 55(6):1867-1882.
7. Andersen et al. (2000) Functional specificity of the replication fork-arrest complexes of Bacillus subtilis and Escherichia coli: significant specificity for Tus-ter functioning in E. coli. Molecular Microbiology. 36(6):1327-1335.
8. Iiamaa, T and Wake, R. (1987) The normal replication terminus of the Bacillus subtilis chromosome, terC, is dispensible for vegetative growth and sporulation. J. Mol. Biol 195:299-310.
9. Roecklein et al. (1991) The tus gene of Escherichia coli: autoregulation, analysis of flanking sequences and identification of a complementary system in Salmonella typhimurium. Res. Microbiol. 142:169-175.
10. Griffiths et al. (2008) Introduction to Genetic Analysis. 9th edition. W H Freeman Publishing, USA.
11. Wilce et al. (2001) Structure of the RTP-DNA complex and the mechanism of polar replication fork arrest. Nature structural biology 8(3):206-210.
12. Duggin et al. (2005) A complex mechanism determines polarity of DNA replication fork arrest by the replication terminator complex of Bacillus subtilis. J. Biol. Chem. 280(13):13105-13113.