SB2013 L04gr5
VlsEVlsE
Lyme disease is the most pervasive tick-borne disease in Europe, the United States,and parts of Asia.[1] It is a multistage infection caused by the spirochete Borrelia burgdorferi. Early symptoms include headaches, depression, rash, and fever. If Lyme disease is left untreated, serious complications of the joints, heart and central nervous system can occur. Infected Ixodes ticks in their nymph stage transmit Lyme disease by attaching to humans and other mammals[2]. In most cases, the tick must be attached for 36-48 hours before the bacteria can be transmitted.[3]
Vmp-like sequence Expressed protein (VlsE), is a lipoprotein on the surface of Borrelia burgdorferi. VlsE contributes to the immune evasion and persistence of Lyme disease.[1] VlsE contains invariable and variable domains. It was found that the varaible domain is highly immunogenic and the target of the immune response; however, through antigenic variation the lipoprotein is able to evade the host immune system.[4]
Within the variable domain, there are invariable regions that remain unchanged during antigenic variation, and therefore may be targets of an immune response. IR6, the most conserved IR, has been found to be immunodominant. The antigencity of each of the 6 IRs has been studied using peptide-based enzyme linked immunosorbent essays, ELISA.[4]
|
StructureStructure
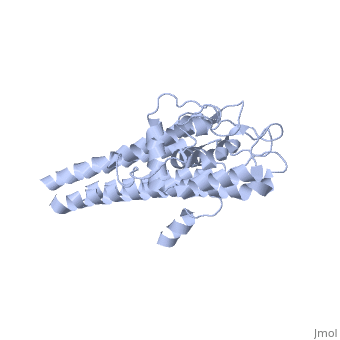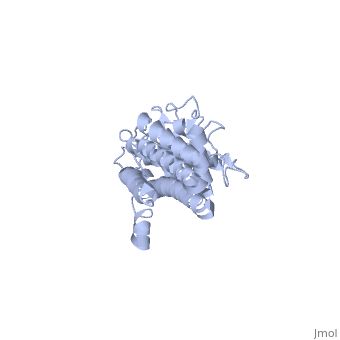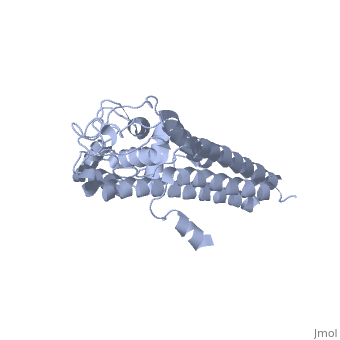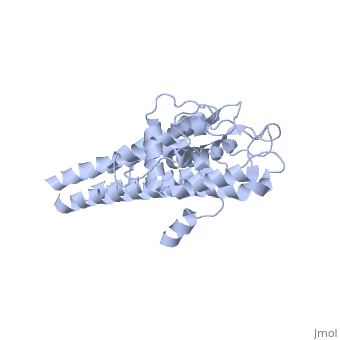
The lipoprotein, VlsE, consists of two invariable domains at the amino and carboxyl termini and a variable domain (Figure 1). When referring to the primary structure of the protein, the variable domain is a cassette region located between the two termini. The variable domain can be further broken down into variable regions () and invariable regions () (Liang et al. 1999).
When crystallized, VlsE forms a four molecule asymmetric unit with each molecule having slight differences in their conformation. Although each molecule in the unit is slightly different, a single molecule of the protein consists of eleven α-helices and four short β-strands. Helices α1 (aa 306-341), α2 (aa 68-87), α3 (aa 114-139), and α11 (aa 306-341) all form the of VlsE, while helices α4 through α10 form the primary region of the of the protein. The four short β-strands each consist of 3 amino acids and can also be located in the membrane distal region.
Covering the membrane distal part of VlsE are connecting loop regions, which lack secondary structure and have different conformations in each of the molecules. (Eicken et al. 2002). Helices α3 through α10 form the invariable regions and are attached by the connecting loops that are classified as the variable regions. Although VlsE crystallizes into an asymmetrical unit, it appears primarily as monomeric in solution. Because the interface between VlsE molecules in the crystal structure buries approximately 13% of the accessible surface area of the monomers, Eicken et al.suggest that there is a possibility of VlsE existing as a dimer when in its natural state (Figure 2).
 Figure 1 shows the different domains of VlsE.
Figure 1 shows the different domains of VlsE.
 Figure 2 shows the possible dimerization of VlsE.
Figure 2 shows the possible dimerization of VlsE.
Antigenic VariationAntigenic Variation
Antigenic variation is the process by which an organism is able to evade its host’s immune system. The antigenic variation that occurs in VlsE uses a complex genetic conversion mechanism that is seen to be concentrated in the VRs. The complex genetic system for VlsE is located near the right telomere on a 28-kb linear plasmid (lp28-1) in strains of B. burgdorferi. This locus has been shown to be a crucial element for the persistence and virulence of Lyme disease (Bankhead and Chaconas 2007). The vls antigenic variation locus consists of a vls expression site (vlsE) and 15 silent vls cassettes that reside just upstream of the site. The vlsE cassette region, which is the variable domain and does not include the invariable amino or carboxyl termini, has approximately 92% DNA sequence identity with the silent vls cassettes. However, the silent vls cassettes lack promoter sequences and are therefore not expressed. Throughout the course of infection, the sequences for the flanking termini and the silent vls cassettes are conserved while the vlsE sequence is recombined. This suggests that the genetic variation mechanism occurs by copying segments of the 15 silent vls cassettes and completely replacing corresponding segments of vlsE sequences. The resulting differences are centralized in the highly variable regions of the vls cassettes. This allows for the constant evolution of the VR structures and evades the antibodies of the host immune system Cite error: Closing </ref> missing for <ref> tag
2.[2]
3.[3]
4.[4]
5.[5]
6.[6]
9.[7]
10.[8]
11.[9]
</ref>
}}