Hemoglobin: Difference between revisions
Eran Hodis (talk | contribs) No edit summary |
Eran Hodis (talk | contribs) |
||
| Line 20: | Line 20: | ||
*{Kin 2}* An exercise in allostery - the Hb tetramer T -> R transition | *{Kin 2}* An exercise in allostery - the Hb tetramer T -> R transition | ||
For hemoglobin, its function as an oxygen-carrier in the blood is fundamentally linked to the equilibrium between the two main states of its quaternary structure, the unliganded "deoxy" or "T state" versus the liganded "oxy" or "R state". The T-state is shown in shades of blue (bluetint alpha-chains, cyan betas, and skyblue hemes) and the R-state is in shades of pink (pinktint alphas, pink betas, and hotpink hemes), suggestive of the change in color between deoxygenated and oxygenated blood. The unliganded (deoxy) form is called the "T" (for "tense") state because it contains extra stabilizing interactions between the subunits. In the high-affinity R-state conformation the interactions which oppose oxygen binding and stabilize the tetramer are somewhat weaker or "relaxed". In some organisms this difference is so pronounced that their Hb molecules dissociate into dimers in the oxygenated form. Animation of the structural changes that occur during this transition can illuminate how such changes result in important functional properties, such as cooperativity of oxygen binding and allosteric control by pH and anions. Hemoglobin is definitely not a pure two-state system, but the T to R transition provides the major, first-level explanation of its function. | |||
The hemoglobin molecule (or "Hb") is a tetramer of two alpha and two beta chains, of 141 and 146 residues in human. They are different but homologous, with a "globin fold" structure similar to myoglobin (Kin. 3.6). The two crystal structures used here are human deoxy hemoglobin (PDB file 3HHB), which is in the T-state quaternary structure with no ligands at the O2 binding sites, and human carbonmonoxy hemoglobin (PDB file 1HCO), which is in the R-state quaternary structure and has ligands at all 4 sites. | |||
Kinemage 1 shows a single alpha chain of hemoglobin, starting with an overview of the subunit. The 6 major and 2 short alpha-helices that make up the structure of a Hb subunit (the "globin fold") are labeled A through H, which is the traditional naming scheme. For example, the proximal histidine (the tightest protein Fe ligand) is often called His F9, since it is residue 9 on helix F (it is residue 87 in the human alpha chain). The helices form an approximately-cylindrical bundle, with the heme and its central Fe atom bound in a hydrophobic pocket between the E and F helices. Turn on "highlights" and click the "animate" button, or press the "a" key on the keyboard, to cycle between the deoxy and oxy forms in this overview. | |||
Click here: *{Kinemage 1, View 2, m= {highlights} on}* for a closeup around the heme O2-binding site. Animate back & forth between the deoxy and oxy forms. For this kinemage the oxy and deoxy alpha1 heme groups were superimposed on each other, to give a local comparison at this site. The heme is quite domed in the blue T-state (deoxy) form, with the 5-coordinate, high-spin Fe (yellow ball) out of the plane. In the pink R-state form a CO molecule is bound at the left; the Fe, now 6-coordinate low-spin, has moved into the heme plane, which has flattenened. The proximal His (at right) connects the Fe to helices on the proximal side, making the Fe position sensitive to changes in the globin structure and vice versa. Remember that this kinemage shows a subunit in the all-unliganded versus the all-liganded states of Hb; when oxygen binds to just one subunit, then its internal structure undergoes some but not all of these changes, depending on conditions. | |||
O2 binds in the same place as CO, with similar effects on the structure; however, for O2 the outer atom is angled rather than straight. The equilibrium between free and bound O2 is very rapid, with on and off rates that are sensitive to protein conformation. Both CO and NO dissociate from the Fe atom very slowly, so that these gases act as respiratory poisons. The alpha and beta chains differ somewhat in their rates and relative affinities for O2 and other ligands, by virtue of heme-pocket differences, but the differences between affinities in the R vs T quaternary states are much larger. | |||
Both alpha and beta chains of Hb resemble myoglobin (the single-chain O2-binder in muscle), both in overall tertiary structure and in using an Fe atom centered in a heme group as the site where oxygen is reversibly bound. The heme is surrounded by a hydrophobic pocket, which is necessary in order for it to bind oxygen reversibly without undergoing oxidation or other undesirable reactions. Click here: *{Kin 1, View 3, m= {Hphobics} on}* to see some of the hydrophobic sidechains that form the heme pocket. They actually surround the binding site so thoroughly that O2 cannot get in or out without parts of the protein moving out of the way a bit, so that its dynamic properties are essential to have any O2 binding at all; this restrictive process also increases the specificity of ligand binding. | |||
The shift between R and T state requires subunit interactions and does not occur in myoglobin, or in isolated alpha or beta chain monomers. These monomers bind O2 quite tightly, which would work well for loading O2 in the lungs but would not allow unloading it for delivery to the tissues. Therefore, the central critical feature of hemoglobin function is how it achieves, uses, and allosterically controls cooperativity between the 4 binding sites in the tetramer to tune O2 binding for satisfying physiological needs. | |||
Click here: *{Kin 1, View 4, m= {Hphobics} off}* and animate, to see the linkage between changes at the O2-binding site and changes in protein conformation, which shows ligand-dependent shifts in the region from the heme out to the subunit interface. Linkage of the heme Fe through the proximal His results in tertiary-structure changes that can then transmit their effects to other subunits in the tetrameric assemblage. This allows O2 binding in one subunit to indirectly affect the affinitiy of other subunits. Briefly, inside the alpha chains the R/T equilibrium is reflected in changes in Fe spin state and position as it moves in or out of the heme plane; the proximal His changes distance and angle relative to the heme; the F helix shifts; Tyr 140 moves and its H-bond to backbone weakens; and both the C-terminus of the chain and Arg 141 move significantly at the interface. Changes at the subunit interface (coupled with changes at the Fe, as we have seen) alter the equilibrium between the deoxy and oxy quaternary structures, and conversely a change of quaternary structure alters the balance between the two states inside a given subunit. Each O2 that binds increases the likelihood of switching the tetramer into the oxy state, and once it switches, the O2 affinity at all sites increases because the local structure changes have either already occurred or are easier to make. | |||
Click here: *{Kin 1, View 5, m= {highlights} off, m= {axes} on}* to show the alpha1 subunit, but centered for the whole tetramer (deoxy form), as it will be seen in View1 of Kinemage 13, including the 2-fold axes of symmetry of the tetramer. | |||
==Acknowledgements== | ==Acknowledgements== | ||
Revision as of 01:19, 20 November 2007
|
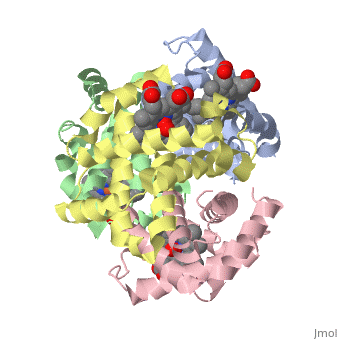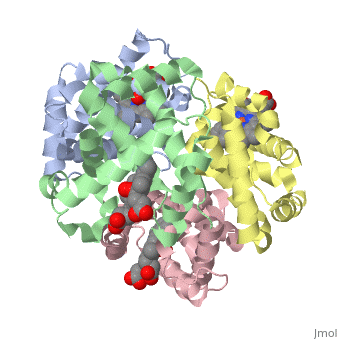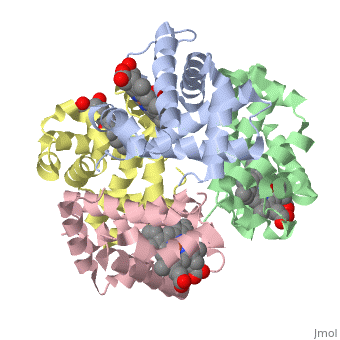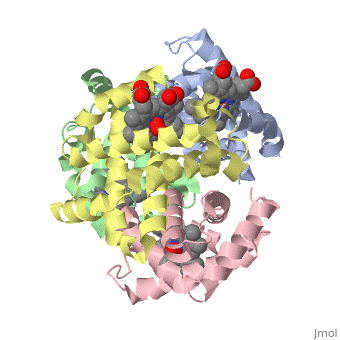
Hemoglobin is an oxygen-transport protein. Hemoglobin is an allosteric protein. It is a tetramer composed of two types of subunits designated α and β, whose stoichiometry is . The of hemoglobin sit roughly at the corners of a tetrahedron, facing each other across a at the center of the molecule. Each of the subunits prosthetic group. The give hemoglobin its red color.
Each individual molecule contains one atom. In the lungs, where oxygen is abundant, an binds to the ferrous iron atom of the heme molecule and is later released in tissues needing oxygen. The heme group binds oxygen while still attached to the . The spacefill view of the hemoglobin polypeptide subunit with an oxygenated heme group shows how the within the polypeptide.
is facilitated by a histidine nitrogen that binds to the iron. A second histidine is near the bound oxygen. The "arms" (propanoate groups) of heme face are hydrophilic and face the surface of the protein while the hydrophobic portions of the heme are buried among the hydrophobic amino acids of the protein. We can also view the anchored heme with the hemoglobin polypeptide subunit shown in a representation.
Secondary structureSecondary structure
Most of the amino acids in hemoglobin form , connected by short non-helical segments. Hemoglobin has no beta strands and no disulfide bonds. A rainbow coloring scheme from N-terminus (blue) to C-terminus (red) helps to discern the . This is at the protein-water interface.
Hydrophobicity, polarity and chargeHydrophobicity, polarity and charge
This view of the shows charged amino acids (lime green), polar amino acids (dark green), hydrophobic amino acids (grey), heme (tan), and water oxygens (light blue). Charged and polar amino acids are common on the surface, where they form hydrogen bonds with water. Most water is not shown. Protein structures tend to fold in such a way so that their hydrophobic residues are buried and their hydrophilic residues are on the surface. A vertical slice through the molecule near the front surface, shows . As we move the slice plane into the molecule, the center is hydrophobic while the surfaces have many polar and charged residues. We can move the slab , and . Near the rear surface, hydrophilic residues are .
SectionSection
Contents of file HbAllo.kin:
- {Kin 1}* An exercise in allostery - a hemoglobin subunit binding O2
- {Kin 2}* An exercise in allostery - the Hb tetramer T -> R transition
For hemoglobin, its function as an oxygen-carrier in the blood is fundamentally linked to the equilibrium between the two main states of its quaternary structure, the unliganded "deoxy" or "T state" versus the liganded "oxy" or "R state". The T-state is shown in shades of blue (bluetint alpha-chains, cyan betas, and skyblue hemes) and the R-state is in shades of pink (pinktint alphas, pink betas, and hotpink hemes), suggestive of the change in color between deoxygenated and oxygenated blood. The unliganded (deoxy) form is called the "T" (for "tense") state because it contains extra stabilizing interactions between the subunits. In the high-affinity R-state conformation the interactions which oppose oxygen binding and stabilize the tetramer are somewhat weaker or "relaxed". In some organisms this difference is so pronounced that their Hb molecules dissociate into dimers in the oxygenated form. Animation of the structural changes that occur during this transition can illuminate how such changes result in important functional properties, such as cooperativity of oxygen binding and allosteric control by pH and anions. Hemoglobin is definitely not a pure two-state system, but the T to R transition provides the major, first-level explanation of its function.
The hemoglobin molecule (or "Hb") is a tetramer of two alpha and two beta chains, of 141 and 146 residues in human. They are different but homologous, with a "globin fold" structure similar to myoglobin (Kin. 3.6). The two crystal structures used here are human deoxy hemoglobin (PDB file 3HHB), which is in the T-state quaternary structure with no ligands at the O2 binding sites, and human carbonmonoxy hemoglobin (PDB file 1HCO), which is in the R-state quaternary structure and has ligands at all 4 sites.
Kinemage 1 shows a single alpha chain of hemoglobin, starting with an overview of the subunit. The 6 major and 2 short alpha-helices that make up the structure of a Hb subunit (the "globin fold") are labeled A through H, which is the traditional naming scheme. For example, the proximal histidine (the tightest protein Fe ligand) is often called His F9, since it is residue 9 on helix F (it is residue 87 in the human alpha chain). The helices form an approximately-cylindrical bundle, with the heme and its central Fe atom bound in a hydrophobic pocket between the E and F helices. Turn on "highlights" and click the "animate" button, or press the "a" key on the keyboard, to cycle between the deoxy and oxy forms in this overview.
Click here: *{Kinemage 1, View 2, m= {highlights} on}* for a closeup around the heme O2-binding site. Animate back & forth between the deoxy and oxy forms. For this kinemage the oxy and deoxy alpha1 heme groups were superimposed on each other, to give a local comparison at this site. The heme is quite domed in the blue T-state (deoxy) form, with the 5-coordinate, high-spin Fe (yellow ball) out of the plane. In the pink R-state form a CO molecule is bound at the left; the Fe, now 6-coordinate low-spin, has moved into the heme plane, which has flattenened. The proximal His (at right) connects the Fe to helices on the proximal side, making the Fe position sensitive to changes in the globin structure and vice versa. Remember that this kinemage shows a subunit in the all-unliganded versus the all-liganded states of Hb; when oxygen binds to just one subunit, then its internal structure undergoes some but not all of these changes, depending on conditions.
O2 binds in the same place as CO, with similar effects on the structure; however, for O2 the outer atom is angled rather than straight. The equilibrium between free and bound O2 is very rapid, with on and off rates that are sensitive to protein conformation. Both CO and NO dissociate from the Fe atom very slowly, so that these gases act as respiratory poisons. The alpha and beta chains differ somewhat in their rates and relative affinities for O2 and other ligands, by virtue of heme-pocket differences, but the differences between affinities in the R vs T quaternary states are much larger.
Both alpha and beta chains of Hb resemble myoglobin (the single-chain O2-binder in muscle), both in overall tertiary structure and in using an Fe atom centered in a heme group as the site where oxygen is reversibly bound. The heme is surrounded by a hydrophobic pocket, which is necessary in order for it to bind oxygen reversibly without undergoing oxidation or other undesirable reactions. Click here: *{Kin 1, View 3, m= {Hphobics} on}* to see some of the hydrophobic sidechains that form the heme pocket. They actually surround the binding site so thoroughly that O2 cannot get in or out without parts of the protein moving out of the way a bit, so that its dynamic properties are essential to have any O2 binding at all; this restrictive process also increases the specificity of ligand binding.
The shift between R and T state requires subunit interactions and does not occur in myoglobin, or in isolated alpha or beta chain monomers. These monomers bind O2 quite tightly, which would work well for loading O2 in the lungs but would not allow unloading it for delivery to the tissues. Therefore, the central critical feature of hemoglobin function is how it achieves, uses, and allosterically controls cooperativity between the 4 binding sites in the tetramer to tune O2 binding for satisfying physiological needs.
Click here: *{Kin 1, View 4, m= {Hphobics} off}* and animate, to see the linkage between changes at the O2-binding site and changes in protein conformation, which shows ligand-dependent shifts in the region from the heme out to the subunit interface. Linkage of the heme Fe through the proximal His results in tertiary-structure changes that can then transmit their effects to other subunits in the tetrameric assemblage. This allows O2 binding in one subunit to indirectly affect the affinitiy of other subunits. Briefly, inside the alpha chains the R/T equilibrium is reflected in changes in Fe spin state and position as it moves in or out of the heme plane; the proximal His changes distance and angle relative to the heme; the F helix shifts; Tyr 140 moves and its H-bond to backbone weakens; and both the C-terminus of the chain and Arg 141 move significantly at the interface. Changes at the subunit interface (coupled with changes at the Fe, as we have seen) alter the equilibrium between the deoxy and oxy quaternary structures, and conversely a change of quaternary structure alters the balance between the two states inside a given subunit. Each O2 that binds increases the likelihood of switching the tetramer into the oxy state, and once it switches, the O2 affinity at all sites increases because the local structure changes have either already occurred or are easier to make.
Click here: *{Kin 1, View 5, m= {highlights} off, m= {axes} on}* to show the alpha1 subunit, but centered for the whole tetramer (deoxy form), as it will be seen in View1 of Kinemage 13, including the 2-fold axes of symmetry of the tetramer.