Histone: Difference between revisions
Michal Harel (talk | contribs) No edit summary |
Ann Taylor (talk | contribs) No edit summary |
||
| Line 1: | Line 1: | ||
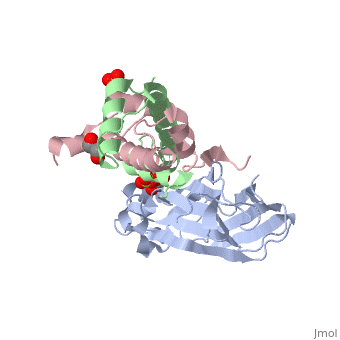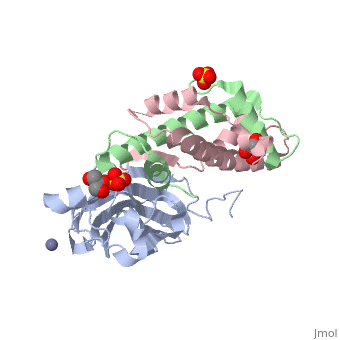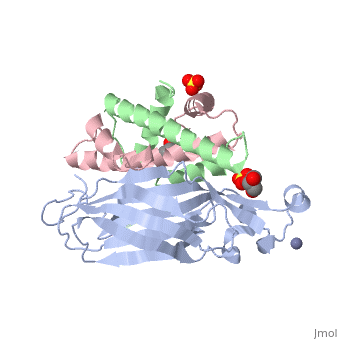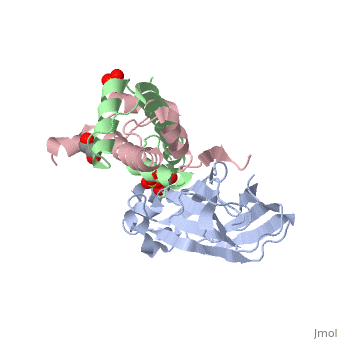
<StructureSection load='2hue' size='340' side='right' caption='Yeast H3 (green), H4 (pink) and anti-silencing protein (grey) complex with sulfate, glycerol and Zn+2 ion (grey), [[2hue]]' scene=''> | <StructureSection load='2hue' size='340' side='right' caption='Yeast H3 (green), H4 (pink) and anti-silencing protein (grey) complex with sulfate, glycerol and Zn+2 ion (grey), [[2hue]]' scene=''> | ||
''' | ==Histone core protein structure== | ||
* | Histones are highly <scene name='Taylor_histone_sandbox/Conservation/1'>conserved proteins</scene> (more purple = more conserved) with <scene name='Taylor_histone_sandbox/Charge_distribution/1'>positive charge</scene> (blue is positive charge, red is negative charge). Because of this positive charge, they interact electrostatically with the negatively charged phosphate groups in DNA. | ||
There are five major classes of histones: H1/H5, H2A, H2B, H3, and H4.<ref name="Bhasin_2006">{{cite journal | author = Bhasin M, Reinherz EL, Reche PA | title = Recognition and classification of histones using support vector machine | journal = J. Comput. Biol. | volume = 13 | issue = 1 | pages = 102–12 | year = 2006 | pmid = 16472024 | doi = 10.1089/cmb.2006.13.102 | url = }}</ref><ref name="Voet, Voet, and Pratt">{{Cite book|surname1= Voet|given1= Donald |surname2= Voet|given2= Judith|surname3= Pratt|given3= Leon A.| year=1988|title=Basic Genetics|publication-place=Boston|publisher=Jones and Bartlett Publishers|isbn=0-86720-090-1}}</ref> Histones <scene name='46/468228/2a/3'>H2A</scene>, <scene name='46/468228/2b/4'>H2B</scene>, <scene name='46/468228/3/3'>H3</scene>, and <scene name='46/468228/H4/1'>H4</scene> are known as the core histones, while histones H1 and H5 are known as the linker histones. | |||
The 4 'core' histones (H2A, H2B, H3 and H4) are relatively similar in structure and are highly conserved through evolution, all featuring a <scene name='Taylor_histone_sandbox/N_c_rainbow/1'>'helix turn helix turn helix' </scene> motif (which allows the easy dimerization). They also share the feature of long 'tails' on one end of the amino acid structure, which are often covalently modified to regulate gene expression. | |||
== Histone interactions with DNA == | |||
Histones are the chief protein components of <scene name='46/468228/Nucleosome/1'>chromatin</scene>, acting as spools around which DNA winds, and play a role in gene regulation. Without histones, the unwound DNA in chromosomes would be very long; each human cell has about 1.8 meters of DNA, but wound on the histones it has about 90 micrometers (0.09 mm) of chromatin, which, when duplicated and condensed during mitosis, result in about 120 micrometers of chromosomes.<ref name="pmid11893489">{{cite journal | author = Redon C, Pilch D, Rogakou E, Sedelnikova O, Newrock K, Bonner W | title = Histone H2A variants H2AX and H2AZ | journal = Curr. Opin. Genet. Dev. | volume = 12 | issue = 2 | pages = 162–9 | year = 2002 | month = April | pmid = 11893489 | doi = 10.1016/S0959-437X(02)00282-4 | url = }}</ref> DNA is wrapped around nucleosomes with approximately 50 base pairs of DNA between subsequent nucleosomes (also referred to as linker DNA). The assembled histones and DNA is called chromatin. During mitosis and meiosis, the condensed chromosomes are assembled through interactions between nucleosomes and other regulatory proteins. | |||
The nucleosome core is formed of two <scene name='46/468228/H2a_h2b_dimer/1'>H2A-H2B dimers</scene> and a <scene name='46/468228/H4_h3_tetramer/1'>H3-H4 tetramer</scene>, forming two nearly <scene name='46/468228/Nucleosome_dimer/1'>symmetrical halves</scene> by tertiary structure.<ref name=pmid9305837/> 147 base pairs of <scene name='46/468228/Dna_wrap_around_histone/1'>DNA wrap</scene> around this core particle 1.65 times in a left-handed super-helical turn.<ref name=pmid9305837>{{cite journal | author = Luger K, Mäder AW, Richmond RK, Sargent DF, Richmond TJ | title = Crystal structure of the nucleosome core particle at 2.8 A resolution | journal = Nature | volume = 389 | issue = 6648 | pages = 251–60 | year = 1997 | month = September | pmid = 9305837 | doi = 10.1038/38444 | url = }} {{PDB|1AOI}}</ref> The linker histone H1 binds the nucleosome and the entry and exit sites of the DNA, thus locking the DNA into place<ref name="isbn0-915274-84-1">{{cite book |author=Farkas, Daniel |title=DNA simplified: the hitchhiker's guide to DNA |publisher=AACC Press |location=Washington, D.C |year=1996 |isbn=0-915274-84-1 }}</ref> and allowing the formation of higher order structure. | |||
In all, histones make five types of interactions with DNA: | |||
*<scene name='Taylor_histone_sandbox/Rockets/1'>Helix-dipoles</scene> from alpha-helices in H2B, H3, and H4 cause a net positive charge to accumulate at the point of interaction with negatively charged phosphate groups on DNA | |||
*Hydrogen bonds between the DNA backbone and the peptide bond in the backbone of histone proteins | |||
*Interactions between the histone and deoxyribose sugars on DNA | |||
*<scene name='46/468228/Tails_charge/1'>salt bridges and hydrogen bonds </scene> between side chains of basic amino acids (especially lysine and arginine) and phosphate oxygens on DNA | |||
*Non-specific minor groove insertions of the H3 and H2B <scene name='46/468228/Tails/2'>N terminal tails</scene> into two minor grooves each on the DNA molecule | |||
In general, genes that are active have less bound histone, while inactive genes are highly associated with histones during interphase. It also appears that the structure of histones has been evolutionarily conserved, as any deleterious mutations would be severely maladaptive. | |||
=== Chromatin regulation === | |||
Histones are subject to post translational modification by enzymes primarily on their N-terminal tails, but also in their globular domains. Such modifications include methylation, acetylation, phosphorylation, SUMOylation, ubiquitination, and ADP-ribosylation. This affects gene expression. The core of the histones H2A, H2B, and H3 can also be modified. Combinations of modifications are thought to constitute a code, the so-called "histone code".<ref name=pmid10638745>{{cite journal |author=Strahl BD, Allis CD |title=The language of covalent histone modifications |journal=Nature |volume=403 |issue=6765 |pages=41–5 |date=Jan 2000 |pmid=10638745 |doi=10.1038/47412}}</ref><ref name=pmid11498575>{{cite journal |author=Jenuwein T, Allis CD |title=Translating the histone code |journal=Science |volume=293 |issue=5532 |pages=1074–80 |date=Aug 2001 |pmid=11498575 |doi=10.1126/science.1063127}}</ref> Histone modifications act in diverse biological processes such as gene regulation, DNA repair, chromosome condensation (in mitosis, spermatogenesis, and meiosis).<ref>{{cite journal |author=Ning Song, Jie Liu, Shucai An, Tomoya Nishino, Yoshitaka Hishikawa and Takehiko Koji |year=2011 |title= Immunohistochemical Analysis of Histone H3 Modifications in Germ Cells during Mouse Spermatogenesis |journal=Acta Histochemica et Cytochemica |volume=44 |issue=4 |doi=10.1267/ahc.11027 |url=http://www.jstage.jst.go.jp/article/ahc/advpub/0/advpub_1107140114/_article |pages=183–90 |pmid=21927517 |pmc=3168764}}</ref> | |||
The common nomenclature of histone modifications is: | |||
*The name of the histone (e.g., H3) | |||
*The single-letter amino acid abbreviation (e.g., K for Lysine) and the amino acid position in the protein | |||
*The type of modification (Me: methyl, P: phosphate, Ac: acetylation, Ub: [[ubiquitin]]) | |||
So H3K4me1 denotes the monomethylation of the 4th residue (a lysine) from the start (i.e., the N-terminal) of the H3 protein. | |||
</StructureSection> | |||
* For nucleosome structure see<br /> | * For nucleosome structure see<br /> | ||
* [[User:Eric Martz/Nucleosomes]]<br /> | * [[User:Eric Martz/Nucleosomes]]<br /> | ||
| Line 11: | Line 48: | ||
[[Histone 3D structures]] | [[Histone 3D structures]] | ||
== References == | == References == | ||
Revision as of 18:04, 25 November 2019
Histone core protein structureHistones are highly (more purple = more conserved) with (blue is positive charge, red is negative charge). Because of this positive charge, they interact electrostatically with the negatively charged phosphate groups in DNA. There are five major classes of histones: H1/H5, H2A, H2B, H3, and H4.[1][2] Histones , , , and are known as the core histones, while histones H1 and H5 are known as the linker histones. The 4 'core' histones (H2A, H2B, H3 and H4) are relatively similar in structure and are highly conserved through evolution, all featuring a motif (which allows the easy dimerization). They also share the feature of long 'tails' on one end of the amino acid structure, which are often covalently modified to regulate gene expression.
Histone interactions with DNAHistones are the chief protein components of , acting as spools around which DNA winds, and play a role in gene regulation. Without histones, the unwound DNA in chromosomes would be very long; each human cell has about 1.8 meters of DNA, but wound on the histones it has about 90 micrometers (0.09 mm) of chromatin, which, when duplicated and condensed during mitosis, result in about 120 micrometers of chromosomes.[3] DNA is wrapped around nucleosomes with approximately 50 base pairs of DNA between subsequent nucleosomes (also referred to as linker DNA). The assembled histones and DNA is called chromatin. During mitosis and meiosis, the condensed chromosomes are assembled through interactions between nucleosomes and other regulatory proteins.
The nucleosome core is formed of two and a , forming two nearly by tertiary structure.[4] 147 base pairs of around this core particle 1.65 times in a left-handed super-helical turn.[4] The linker histone H1 binds the nucleosome and the entry and exit sites of the DNA, thus locking the DNA into place[5] and allowing the formation of higher order structure.
In all, histones make five types of interactions with DNA:
In general, genes that are active have less bound histone, while inactive genes are highly associated with histones during interphase. It also appears that the structure of histones has been evolutionarily conserved, as any deleterious mutations would be severely maladaptive. Chromatin regulationHistones are subject to post translational modification by enzymes primarily on their N-terminal tails, but also in their globular domains. Such modifications include methylation, acetylation, phosphorylation, SUMOylation, ubiquitination, and ADP-ribosylation. This affects gene expression. The core of the histones H2A, H2B, and H3 can also be modified. Combinations of modifications are thought to constitute a code, the so-called "histone code".[6][7] Histone modifications act in diverse biological processes such as gene regulation, DNA repair, chromosome condensation (in mitosis, spermatogenesis, and meiosis).[8] The common nomenclature of histone modifications is:
So H3K4me1 denotes the monomethylation of the 4th residue (a lysine) from the start (i.e., the N-terminal) of the H3 protein.
|
| ||||||||||
- For nucleosome structure see
- User:Eric Martz/Nucleosomes
- Nucleosome structure
- Nucleosomes
- Nucleosome structure (Spanish)
3D Structures of histone3D Structures of histone
ReferencesReferences
Cite error: <ref> tag with name "Armstrong" defined in <references> is not used in prior text.