Molecular Playground/DnaK: Difference between revisions
No edit summary |
Michal Harel (talk | contribs) No edit summary |
||
| (50 intermediate revisions by 2 users not shown) | |||
| Line 1: | Line 1: | ||
<applet load=' | <applet load='2kho' size='400' color='white' frame='true' align='right' caption='The E. coli Hsp70, DnaK [[2kho]]'/> | ||
One of the [ | One of the [http://proteopedia.org/wiki/index.php/CBI_Molecules CBI Molecules] being studied in the [http://www.umass.edu/cbi/ University of Massachusetts Amherst Chemistry-Biology Interface Program] at UMass Amherst and on display at the [http://www.molecularplayground.org/ Molecular Playground]. | ||
Molecular Playground banner: DnaK, a central hub in maintaining proteostasis in E. coli | Molecular Playground banner: DnaK, a central hub in maintaining proteostasis in <i>E. coli</i> | ||
<scene name='60/609794/Adp-dnak_1/ | <scene name='60/609794/Adp-dnak_1/4'>DnaK in the extended conformation</scene> | ||
The E. coli Hsp70, DnaK, is a | The <i>E. coli</i> '''Hsp70''', '''DnaK''', is a protein chaperone whose function is to bind exposed hydrohpobic residues of unfolded proteins. This binding event prevents aggregation and rescues the nascent chain from kinetic traps along the folding pathway. Hsp70 protein chaperones switch between an <scene name='60/609794/Atp-dnak/1'>ATP-bound</scene>, low-substrate affinity form and an <scene name='60/609794/Adp-dnak_1/4'>ADP-bound</scene>, high substrate affinity form during their allosteric cycle.[http://www.ncbi.nlm.nih.gov/pubmed/19439666?dopt=Abstract 1] Hsp70 protein chaperones are ubiquitously found in almost all known organisms and cell types and represent a potential target for anti-cancer and neurodegenerative therapies.[http://www.ncbi.nlm.nih.gov/pubmed/21403392 2] | ||
===Structure=== | ===Structure=== | ||
DnaK is a 638 residue protein of approximately 70 kDa. The protein can be thought to be made up of two domains, the N-terminal nucleotide-binding domain (<scene name='60/609794/Nbd/4'>NBD</scene>) (residues 1-388), the C-terminal substrate-binding domain (<scene name='60/609794/Sbd/ | DnaK is a 638 residue protein of approximately 70 kDa. The protein can be thought to be made up of two domains, the N-terminal nucleotide-binding domain (<scene name='60/609794/Nbd/4'>NBD</scene>) (residues 1-388), the C-terminal substrate-binding domain (<scene name='60/609794/Sbd/2'>SBD</scene>) (residues 393-638), which are divided by the <scene name='60/609794/Linker/2'>interdomain linker</scene> (residues 389-392, shown in violet). The NBD is further divided into four subdomains: <scene name='60/609794/Subdomain_ia/2'>IA</scene> (residues 1-37, 112-184, 363-383, shown in red), <scene name='60/609794/Subdomain_ib/2'>IB</scene> (residues 38-111, shown in green), <scene name='60/609794/Subdomain_iia/2'>IIA</scene> (residues 185-227, 310-362, shown in gold), and <scene name='60/609794/Subdomain_iib/2'>IIB</scene> (residues 228-309, shown in purple). The SBD of DnaK consists of a <scene name='60/609794/Beta_basket/1'>beta-basket</scene> (residues 393-507), an <scene name='60/609794/Helical_lid/1'>alpha-helical lid</scene> (residues 508-605), and a disordered C-terminal tail (not shown due to lack of available crystal structure).[http://www.ncbi.nlm.nih.gov/pubmed/22576356 3] | ||
===Function=== | ===Function=== | ||
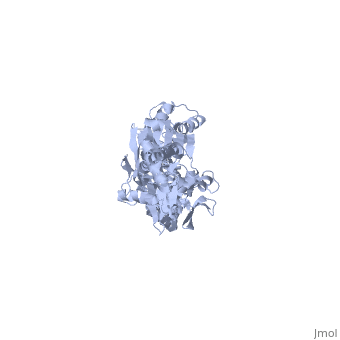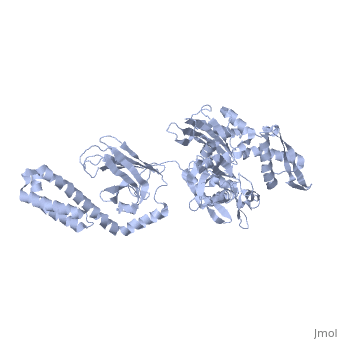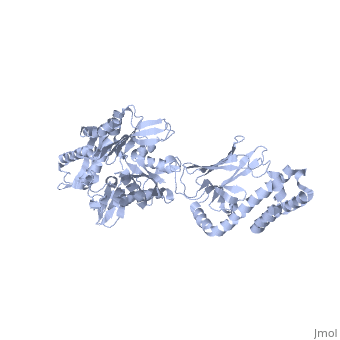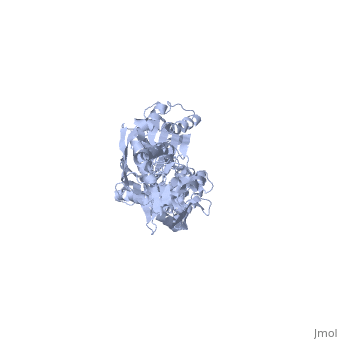
DnaK binds stretches (7-8 residues in length) of exposed hydrophobic residues of its client proteins in order to prevent their aggregation.[http://www.ncbi.nlm.nih.gov/pubmed/14740253] Upon ATP binding, the NBD subdomains rotate relative to each other and induce a conformational change in the NBD (compare the <scene name='60/609794/Subdomain_iib/1'>ADP-bound</scene> NBD to the <scene name='60/609794/Nbd-atp/ | DnaK binds stretches (7-8 residues in length) of exposed hydrophobic residues of its client proteins, in the beta-basket of the SBD, in order to prevent their aggregation.[http://www.ncbi.nlm.nih.gov/pubmed/14740253 4] Upon ATP binding, the NBD subdomains rotate relative to each other and induce a conformational change in the NBD (compare the <scene name='60/609794/Subdomain_iib/1'>ADP-bound</scene> NBD to the <scene name='60/609794/Nbd-atp/2'>ATP-bound</scene> form in <b>Figure 1.</b>[http://www.ncbi.nlm.nih.gov/pubmed/21482798 5] [[Image:Superimposed_nbd.png|right|300x300px|thumb|<b>Figure 1.</b> A superimposition of the ATP-bound and ADP-bound NBDs. The ATP-bound form is shown in lighter colors.]]When ATP binds the NBD, the interdomain linker communicates the allosteric signal to the SBD that induces a conformational change in the SBD, causing the alpha-helical lid to dock onto the NBD.[http://www.ncbi.nlm.nih.gov/pubmed/17434124?dopt=Abstract 6] Compare the <scene name='60/609794/Sbd_open/2'>open</scene> form of the SBD (when the NBD binds ATP) and the <scene name='60/609794/Helical_lid/1'>closed</scene> form of the SBD (when the NBD binds ADP) in <b>Figure 2.</b>[[Image:Superimposed_sbd_3.png|right|300x300px|thumb|<b>Figure 2.</b> A superimposition of the 'open' and 'closed' forms of the SBD. The 'open' form is shown in lighter colors.]] The alpha-helical lid of the SBD rotates 180 degrees during the closing motion.[http://www.ncbi.nlm.nih.gov/pubmed/23123194?dopt=Abstract 7] The result is that the same residues that dock with the NBD are responsible for binding the nascent chain of the client protein. The SBD has low substrate affinity in the 'open' conformation, but high substrate affinity in the 'closed' conformation.[http://www.ncbi.nlm.nih.gov/pubmed/24012426 8] Upon ATP hydrolysis, the NBD reverts to the ADP-bound state, undocking the alpha-helical lid from the NBD and regenerating the 'closed' conformation of the SBD. Alternating cycles of binding and release allow DnaK to unfold kinetically trapped intermediates and allow the client protein to refold to its native state.[http://www.ncbi.nlm.nih.gov/pubmed/20953191 9] Recently, a third 'allosterically active' state of DnaK has been discovered. In this state, the two domains remain undocked, with the SBD retaining high substrate affinity, but the interdomain linker is bound to the NBD. This state occurs when both ATP and substrate are bound simultaneously and has not yet been crystallized.[http://www.ncbi.nlm.nih.gov/pubmed/23217711 10] | ||
===Drug Target=== | ===Drug Target=== | ||
Since | Since Hsp70s are critically positioned in the proteostasis systems of many organisms, including humans, they represent a tempting target for anti-cancer and neurodegenerative disease therapies. Recent efforts to develop a competitive inhibitor for Hsp70s, using DnaK as a model system, have met with only marginal success.[http://www.ncbi.nlm.nih.gov/pubmed/24312699 11] Recently, interactions between Hsp70s and its co-chaperones have been targeted for drug development, but with no leads currently in clinical trials.[http://www.ncbi.nlm.nih.gov/pubmed/22920901 12] | ||
===3D structures of Hsp70=== | ===3D structures of Hsp70=== | ||
[[Heat Shock Proteins]] | |||
===See Also=== | ===See Also=== | ||
The Wikipedia page on Hsp70 is | The Wikipedia page on Hsp70 is useful for a general background.[http://en.wikipedia.org/wiki/Hsp70 14] | ||
This informational video | This informational [https://www.youtube.com/watch?v=yQ3QUDj1Mv8 video] demonstrates the successful folding of a protein in <i>E. coli</i> | ||
The Gierasch lab, in collaboration with the Powers lab at Scripps, has developed a computational model of the proteostasis network in <i>E. coli</i> called FoldEco.[http://www.ncbi.nlm.nih.gov/pubmed/22509487 15] A web version of FoldEco can be found [http://foldeco.scripps.edu here] | |||
===References=== | ===References=== | ||
1. | 1. [http://www.ncbi.nlm.nih.gov/pubmed/19439666?dopt=Abstract Bertelsen EB. et al. Proc Natl Acad Sci USA 2009] | ||
2. [http://www.ncbi.nlm.nih.gov/pubmed/21403392 Broer L. et al. J Alzherimers Dis 2011] | |||
3. [http://www.ncbi.nlm.nih.gov/pubmed/22576356 Zuiderweg ER. et al. Top Curr Chem 2013] | |||
4. [http://www.ncbi.nlm.nih.gov/pubmed/14740253 Wegele H. et al. Rev Physiol Biochem Pharmacol 2004] | |||
5. [http://www.ncbi.nlm.nih.gov/pubmed/21482798 Zhuravleva A. et al. Proc Natl Acad Sci USA 2011] | |||
6. [http://www.ncbi.nlm.nih.gov/pubmed/17434124?dopt=Abstract Swain JF. et al. Mol Cell 2007] | |||
7. [http://www.ncbi.nlm.nih.gov/pubmed/23123194?dopt=Abstract Kityk, R. et al. Mol Cell 2012] | |||
8. [http://www.ncbi.nlm.nih.gov/pubmed/24012426 Mayer MP. Trends Biochem Sci 2013] | |||
9. [http://www.ncbi.nlm.nih.gov/pubmed/20953191 Sharma SK. et al. Nat Chem Bio 2010] | |||
10. [http://www.ncbi.nlm.nih.gov/pubmed/23217711 Zhuravleva A. et al. Cell 2012] | |||
11. [http://www.ncbi.nlm.nih.gov/pubmed/24312699 Li X. et al. ACS Med Chem Lett 2013] | |||
12. [http://www.ncbi.nlm.nih.gov/pubmed/22920901 Assimon VA. et al. Curr Pharm Des 2013] | |||
15. [http://www.ncbi.nlm.nih.gov/pubmed/22509487 Powers ET. et al. Cell Rep 2012] | |||
Latest revision as of 16:23, 6 January 2016
|
One of the CBI Molecules being studied in the University of Massachusetts Amherst Chemistry-Biology Interface Program at UMass Amherst and on display at the Molecular Playground.
Molecular Playground banner: DnaK, a central hub in maintaining proteostasis in E. coli
The E. coli Hsp70, DnaK, is a protein chaperone whose function is to bind exposed hydrohpobic residues of unfolded proteins. This binding event prevents aggregation and rescues the nascent chain from kinetic traps along the folding pathway. Hsp70 protein chaperones switch between an , low-substrate affinity form and an , high substrate affinity form during their allosteric cycle.1 Hsp70 protein chaperones are ubiquitously found in almost all known organisms and cell types and represent a potential target for anti-cancer and neurodegenerative therapies.2
StructureStructure
DnaK is a 638 residue protein of approximately 70 kDa. The protein can be thought to be made up of two domains, the N-terminal nucleotide-binding domain () (residues 1-388), the C-terminal substrate-binding domain () (residues 393-638), which are divided by the (residues 389-392, shown in violet). The NBD is further divided into four subdomains: (residues 1-37, 112-184, 363-383, shown in red), (residues 38-111, shown in green), (residues 185-227, 310-362, shown in gold), and (residues 228-309, shown in purple). The SBD of DnaK consists of a (residues 393-507), an (residues 508-605), and a disordered C-terminal tail (not shown due to lack of available crystal structure).3
FunctionFunction
DnaK binds stretches (7-8 residues in length) of exposed hydrophobic residues of its client proteins, in the beta-basket of the SBD, in order to prevent their aggregation.4 Upon ATP binding, the NBD subdomains rotate relative to each other and induce a conformational change in the NBD (compare the NBD to the form in Figure 1.5

When ATP binds the NBD, the interdomain linker communicates the allosteric signal to the SBD that induces a conformational change in the SBD, causing the alpha-helical lid to dock onto the NBD.6 Compare the form of the SBD (when the NBD binds ATP) and the form of the SBD (when the NBD binds ADP) in Figure 2.

The alpha-helical lid of the SBD rotates 180 degrees during the closing motion.7 The result is that the same residues that dock with the NBD are responsible for binding the nascent chain of the client protein. The SBD has low substrate affinity in the 'open' conformation, but high substrate affinity in the 'closed' conformation.8 Upon ATP hydrolysis, the NBD reverts to the ADP-bound state, undocking the alpha-helical lid from the NBD and regenerating the 'closed' conformation of the SBD. Alternating cycles of binding and release allow DnaK to unfold kinetically trapped intermediates and allow the client protein to refold to its native state.9 Recently, a third 'allosterically active' state of DnaK has been discovered. In this state, the two domains remain undocked, with the SBD retaining high substrate affinity, but the interdomain linker is bound to the NBD. This state occurs when both ATP and substrate are bound simultaneously and has not yet been crystallized.10
Drug TargetDrug Target
Since Hsp70s are critically positioned in the proteostasis systems of many organisms, including humans, they represent a tempting target for anti-cancer and neurodegenerative disease therapies. Recent efforts to develop a competitive inhibitor for Hsp70s, using DnaK as a model system, have met with only marginal success.11 Recently, interactions between Hsp70s and its co-chaperones have been targeted for drug development, but with no leads currently in clinical trials.12
3D structures of Hsp703D structures of Hsp70
See AlsoSee Also
The Wikipedia page on Hsp70 is useful for a general background.14
This informational video demonstrates the successful folding of a protein in E. coli
The Gierasch lab, in collaboration with the Powers lab at Scripps, has developed a computational model of the proteostasis network in E. coli called FoldEco.15 A web version of FoldEco can be found here
ReferencesReferences
1. Bertelsen EB. et al. Proc Natl Acad Sci USA 2009
2. Broer L. et al. J Alzherimers Dis 2011
3. Zuiderweg ER. et al. Top Curr Chem 2013
4. Wegele H. et al. Rev Physiol Biochem Pharmacol 2004
5. Zhuravleva A. et al. Proc Natl Acad Sci USA 2011
6. Swain JF. et al. Mol Cell 2007
7. Kityk, R. et al. Mol Cell 2012
8. Mayer MP. Trends Biochem Sci 2013
9. Sharma SK. et al. Nat Chem Bio 2010
10. Zhuravleva A. et al. Cell 2012
11. Li X. et al. ACS Med Chem Lett 2013