Sandbox CYP450
Drug Metabolizing Cytochrome P450 Enzymes and their inhibitorsDrug Metabolizing Cytochrome P450 Enzymes and their inhibitors
This tutorial covers the basic structure and function of the Cytochrome P450 (CYP or CYP450) enzymes, with particular emphasis on their relevance to drug action. It may be useful as a standalone primer on the basics of CYP metabolism, but is intended to supplement a more comprehensive text, such as Chapter 4 of the 7th edition of Foye's Principles of Medicinal Chemistry[1]. The primary purpose of this page is to graphically illustrate some of the principles of CYP450 metabolism that are otherwise difficult to visualize.
CYP450 enzymes are the most important group of drug metabolizing enzymes. A very large proportion of medications are metabolized primarily through oxidation by various CYP450s. There are more than 50 individual CYP450s, but six are quite significant, as they can metabolize from 75-90% of all drugs [2]. These six include: CYP1A2, CYP2C9, CYP2D6, CYP2C19, CYP2D6, CYP3A4 and CYP3A5. Other isoforms such as CYP2E1 and CYP1A1 are important to a lesser degree, such as in the context of activation of procarcinogens. CYP enzymes in families 5 or higher are typically important for processing steroids in humans, rather than drug metabolism. These enzymes are generally known by names other than their CYP designation. For instance, CYP27A1 is known as sterol 27-hydroxylase, and is important for biosynthesis of bile acids. Other CYPs are not found in humans, but may be drug targets. One of these is CYP51A1, or lanosterol 14 alpha-demethylase. This CYP is important for the production of ergosterol in fungi, which serves the same purpose in fungal cell membranes as cholesterol does in humans.
The name of a CYP450 enzyme indicates its similarity in structure to other CYPs. Those with greater than 40% amino acid homology are grouped into families, and those with greater than 55% homology are grouped into subfamilies. There is a systematic method for naming and grouping them, with the letters and numbers referring to families, subfamilies, and individual members. Thus CYP1, CYP2, and CYP3 are separate families. A letter following the family name indicates the particular subfamily. Thus CYP1A1 and CYP1A2 are the first and second members of the CYP1A subfamily, but both belong to the CYP1 family. Individual CYPs, such as CYP1A1, CYP1A2, CYP3A4, and CYP2D6, are referred to as isoforms of each other.
Recognizing the individual CYP responsible for metabolism of a drug can be very important for predicting potentially dangerous results from its co-administration with other drugs. There are several reasons that drugs can have negative interactions based on CYP enzymes. If one drug induces or inhibits a CYP, then another drug metabolized by that same CYP may have increased or decreased levels in the plasma. The result of this might be either toxicity or treatment failure. Interactions are most clinically important in drugs that have a narrow therapeutic range, those that are vital for maintaining a healthy state, and those with important toxicities.
As you go through this tutorial, try to answer each of the questions posed within the text. When you get to the end, write out the answers to the Focused Questions.
Interacting with the Molecular DisplayIn this tutorial, the blue links are standard hyperlinks. The green links show you a particular view, or scene, of the molecule in the interactive window to the right. As you go through the text, click on the green links to show the structural features being highlighted. The first example illustrated here is the second protein discovered in the CYP 1 family, in subfamily A (generally referred to as CYP1A2). This protein is shown in an interactive window to the right, and comes from the PDB entry 2hi4. Turn off/on (toggle) spinning of the protein by clicking on the button below the structure. The quality of the molecule image can also be increased by clicking the "toggle quality" button, although displaying it this way may decrease the smoothness when the molecule is rotating. Now rotate the molecule by clicking and dragging in the window with your cursor or using the scroll wheel on your mouse. Re-size the molecule by holding down the shift key and dragging up and down. Rotate and re-size the molecule until you can clearly see that there are 2 molecules shown in a "space-filling" representation in the middle of the protein (they are almost perpendicular to each other, and almost touching). These are a heme molecule, which is absolutely vital for the enzyme's function, and a second molecule (alpha-naphthoflavone) which is a compound about to be metabolized. As you go through this tutorial, rotate and re-size the molecules as necessary to see the concepts being illustrated. You might also find it useful to toggle the spin or quality of the display.
The Heme Group is Vital for the Enzyme's ActionLets look at the heme now. Click the following link to hide the rest of the protein and . Carbon atoms are shown in grey, Nitrogen atoms are blue, Oxygen is red, and the central Iron atom is orange. The Iron atom is a vital center for oxidation of substrates (drugs or other xenobiotics). Rotate and re-size as necessary to shown that the heme is highly planar. Do you recognize the 4 pyrrole rings within the heme? Examine the Iron atom in the heme. It is capable of forming 6 bonds. Four of those are with the 4 pyrrole rings that make up the porphyrin ring system. The fifth and sixth bonds are made to atoms above and below the plane of heme ring. The fifth is made to a cysteine residue present on the protein. The final bond is made to an Oxygen molecule (not shown). This Oxygen is activated for oxidation of substrates, and would appear directly between the substrate and the Iron atom. The cysteine residue that coordinates the Iron atom is vital, since it helps polarize it for binding the Oxygen. This residue is highly conserved throughout all the known CYP450s. The bond between the Sulfur atom of the cysteine residue is shown (in yellow) . Note that the sixth site for a bond on the Iron is unfilled, and ready to bind to Oxygen. Although the presence of the cysteine residue is invariable in all CYPs, the other residues that surround it can vary, changing the shape of the active site and the drugs that can bind there. Molecules Can be Oxidized at More than One PositionNow look at the substrate molecule being oxidized and view its orientation relative to the heme group by clicking . Re-size and rotate the molecules until you can see how the two molecules are oriented in relationship to each other. The flavone is metabolized (oxidized) by introduction of a hydroxy group onto the phenyl ring attached to the tricyclic ring system. Note particularly the distance and orientation of the heme Iron and the phenyl substituent. When some drugs are administered to humans, the metabolites observed may be the result of oxidation at more than one position in the molecule. What factors do you think might contribute to this? Several factors may come into play. First, more than one CYP isoform may be contributing, with each isoform preferring different sites of attack. Considering only one CYP, like that shown here, there is also the possibility that attack might occur at more than one position, giving more than one metabolite. What factors do you think might lead to this? In this example, purely based on the distance between the phenyl ring of the flavone and the heme iron, oxidation might be expected to take place at the 4 (para) position. However, other factors may also help determine the regioselectivity of the metabolism (selectivity for oxidation at the different possible positions). One of these factors is the relative reactivity of the various positions on the substrate.
Different CYPs can be Selective for Different DrugsDifferent CYPs have preferences for oxidizing different substrates. Some of them have a much narrower range of molecular structures that they can oxidize. CYP1A2 preferentially oxidizes molecules that are relatively large and planar, as in this structure [3]. The complementary size and shape of the drug and binding site makes them a good match for each other. The substrate (flavone) shown here is different from many other substrates of this CYP. Besides being a substrate of CYP1A2, it is also a competitive inhibitor[4]. It can inhibit the metabolism of other CYP1A2 substrates because it binds very tightly to it. Thus, while the CYP is inhibited, it is unavailable to metabolize other substrates (e.g. drugs). Therefore, if a second drug that is a CYP1A2 substrate is co-administered with this compound, it might not be broken down as expected, and could build up to toxic levels. The reason that this flavone is bound so tightly to the enzyme is that its shape and electronic charges are complementary to the binding pocket. This is examined next.
Amino Acids in the Binding Pocket Affect Drug SelectivityFirst examine the shape of the Van der Waals area The green to orange range of colors on the surface show places where there are close contacts with the binding pocket. These contacts can be caused by ionic, hydrophobic, or hydrogen bonding. Blue areas show where the interaction distance is not as close. Given the structure of the flavone, what forces do you suspect may be responsible for binding the enzyme so tightly? In the next scene, the Van der Waals surface of the cavity is . The flavone is now seen as in orange, and portions of the binding pocket most important for binding are shown as patches. These result from specific amino acid residues that form the surface of the binding pocket. Now let's examine the amino acid residues that form that contact surface. Clicking on this will show the surface of the flavone and protein atoms that are within a few angstroms of the inhibitor. The cavity surface is shown translucent to help see what atoms are involved. You can toggle between showing the ligand alone and the ligand with the protein residues by clicking the link again. The red sphere in this view is the oxygen atom of a water molecule inside the active site of the protein. In some cases, water molecules are necessary to help bind substrates to enzymes. What is the interaction between the flavone and this water molecule? In order to put the flavone-protein binding in more context, you can show additional residues that help form the binding pocket. This link can also be toggled on and off. As you rotate the molecule, look at how each of the amino acid residues at the active site is interacting with parts of the flavone. Can you predict what kinds of interactions (H-bonds, lipophilic, or ionic) might be made? Examine the hydrophobic portions of the inhibitor. What kind of residues would interact with these? Hover the cursor over each residue and examine the label that appears (you may have to re-size the molecule to pick the individual atoms). Is the label consistent with your expectations for these kinds of interactions? Since the CYPs largely metabolize hydrophobic substrates, hydrophobic interactions are very important for binding many substrates. That is clearly the case here. The inhibitor shown here can fits exceptionally well in the binding pocket due to the principle of induced fit. Induced fit occurs when a drug binds to a protein and causes a conformational change that leads to tighter binding. Thus the "fit" of the drug to the protein is induced in many cases. A molecule that is able to induce a fit in a protein might be expected to be a competitive inhibitor. Using molecular visualizations such as we are using here can give important insights into the pharmacokinetic behavior of some drugs. Some of these will be examined in the next section.
CYP450 3A4 is Very Important for Drug MetabolismThe CYP3A4 isoform, PDB entry 1tqn, is involved in metabolizing 50% or more of marketed drugs, and metabolizes more drugs than all the other isoforms combined. It is also the enzyme most commonly associated with undesired drug-drug interactions. CYP3A4 differs from the CYP1A2 isoform considered above, as well as most other CYPs, because of the diversity of drugs it can metabolize. A number of these drugs are known to cause potentially dangerous or even fatal interactions. The reason that CYP3A4 is involved in metabolism of so many drugs appears to be related to the size and flexibility of its binding pocket, which can accommodate a number of fairly large drugs. These drugs include the macrolide antibiotic erythromycin, the anticancer drug taxol, the immunosuppresant cyclosporine, and several statins. Examination of drugs such as those bound to CYP3A4 gives some insight into its selectivity and the way that drug binding may influence pharmacokinetics. In the crystal structure of CYP3A4 bound to erythromycin, the volume of the active site is notably different than the unbound protein. Erythromycin is a fairly large drug (MW 733.9), and CYP450 isoforms with smaller binding pockets (e.g. CYP1A2) cannot accommodate it and therefore cannot metabolize it. Due to the large size of the binding pocket, CYP3A4 can also accommodate a diversity of other, smaller substrates.
Effect of Drug Binding on a CYP450 Active SiteThe following scene shows free from any substrate or inhibitor. This may take a moment to load. The surface of the binding pocket is shown in orange and its translucency can be toggled by clicking the link again. Examine the size and shape of the cavity by rotating and resizing the molecule. Observe that the cavity extends within a short distance of the heme ring. In order for a drug to be oxidized, it must come quite close to the oxygen atom held by the heme. Rotate and re-size the molecule until you can clearly see that there are 2 regions open to the outside of the molecule. Note that the opening to the cavity does not seem large enough to allow a drug of any significant size to enter. In order for a larger drug to enter the inside of the enzyme, some change in shape must occur. Next, look at this scene showing CYP3A4 bound to erythromycin, with the of its binding pocket shown in green (PDB entry 2j0d). You may click the link again to toggle the translucency of the isosurface. In this case, CYP3A4 has undergone a conformational change as a result of binding to erythromycin. Note the size and shape of the cavity, and the apparent size of the opening. In comparing the bound and unbound structures, it looks as though the enzyme has adopted a conformation that allows the drug to bind more tightly than its initial interaction. This, again, is induced fit. Finally, looking at both of the structures on each other allows comparison of the active sites before and after binding. After binding, a portion of the erythromycin molecule, shown in ball and stick, is bumping up against a in the protein that was not there prior to binding. This indicates that the drug would not have been able to bind in its current location without induced fit taking place.
Complex Drug Interactions with CYP450sThere are multiple types of inhibitors of CYPs. Competitive inhibition, as seems to be happening with many substrates, follows fairly predictable rules of dose to rate of metabolism. In this case, 2 drugs are competing for the same active site. Some drugs metabolized by CYP3A4, however are known to have more complex interactions. For instance, some substrates can change the apparent binding affinity for other inhibitors and substrates. Other substrates fail to inhibit some specific substrates[5]. The observed effect here is that both drugs are metabolized without interference with each other, which is seemingly a counterintuitive result. It has been demonstrated that 2 or more smaller molecules may bind within an active site at the same time. In this case, drug metabolism can, strangely enough, actually be increased. It has been proposed that when only one molecule of a smaller drug is bound to the active site, that it may have less catalytic efficiency due to the drug being held only loosely inside the large cavity. In this case, it might not be held in an optimum orientation near the heme Iron. However, when two molecules of a smaller drug bind at the same time, one might then be held closer to the activated heme, improving catalytic efficiency. The principle that more than one drug molecule at a time binding to a CYP may influence its pharmacokinetics is illustrated by the following structure of ketoconazole bound to a CYP enzyme (2j0c)[6]. Ketoconazole is an anti-fungal drug that can have unusual pharmacokinetics, in that its apparent plasma concentration does not follow entirely predictable values in relation to its dose. In the structure shown next, of ketoconazole are bound to the CYP. You can see this interaction better by if we the protein. Now you can the visibility of the surface to see the orientation of the molecules in the pocket. One of the ketoconazole molecules is bound directly to the heme ring (see below), while the second molecule has taken up residence in the pocket and appears to be holding the first one in place. The unusual pharmacokinetics of ketoconazole might be explained by the fact that as its concentration increases in the plasma, the activity of the enzyme changes since it is now binding more than 1 molecule of the drug. Irreversible inhibition of CYP450sWhen we examined CYP1A2 above, the flavone inhibited the enzyme simply by virtue of having such complementary features to the binding site that it bound so tightly that it prevented other drugs binding to it. Another way that a drug can inhibit CYP450 enzymes is by formation of a covalent bond that deactivates the active site. This happens with the well-known inhibitor of CYP3A4, . Ritonavir is a HIV protease inhibitor routinely prescribed in combination with other antivirals. It's efficacy as part of a drug "cocktail" stems from the fact that it is a potent irreversible inhibitor of CYP3A4. Irreversible inhibition differs from most cases of competitive inhibition in that the enzyme is permanently deactivated, and must be re-synthesized by the cell. In this case, we have taken advantage of a the inhibition of CYP3A4 to prevent it from metabolizing the other antivirals that it is co-administered with. As you might expect, extreme caution must be taken to prevent unwanted toxicity from other drugs that also may be administered at the same time. This is relatively easy to control when one pharmacist is dispensing all of the medications that a patient may be taking that could otherwise be dangerous. Examine the orientation of ritonavir relative to the . Now let's look at the binding pocket. Look at the complementary shape of the drug and the (click again to toggle the translucency of the isosurface). In this case, in contrast to erythromycin, there has been a covalent bond formed between the thiazole ring of the drug and the heme ring. Do you see how the thiazole ring has to the heme group? In the next section below, we will examine another factor that causes one drug to be metabolized by one CYP, while another might be metabolized by a second one.
|
| ||||||||||
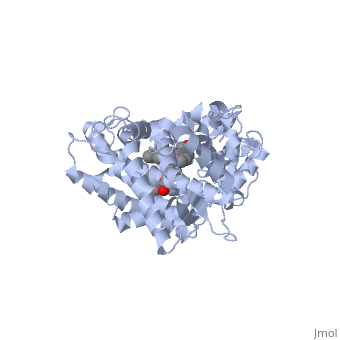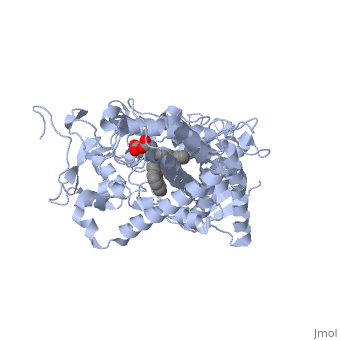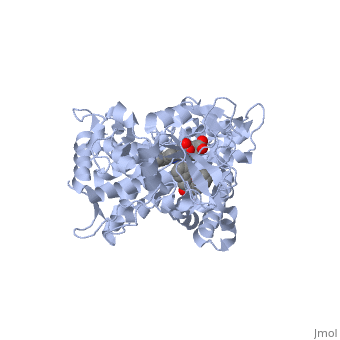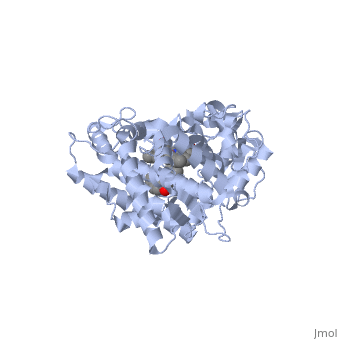
Active site Volume Affects Drug SelectivityActive site Volume Affects Drug Selectivity
Some of the factors that determine which particular CYP450 isoform metabolizes a given drug is the shape and size of its active site. As we saw above, induced fit can cause the shape of a binding pocket to change. However, induced fit may not be enough to allow a CYP with a small binding pocket to open up enough to allow larger drugs to bind. The windows below show two CYP450 isoforms with very different selectivity. The first is the structure of CYP2E1, bound to the enzyme inhibitor 4-methylpyrazole (PDB code 3e4e). The second is the structure of CYP3A4 bound to the inhibitor erythromycin (2j0d). Make sure that the check box below the second window is checked. Now rotate and re-size the structures until you can see the heme group and the size and shape of the active site.
Use one of the buttons below each applet to toggle the transparency of the active site in the CYP2E1 structure and the CYP3A4 structure until you can see how the substrate is bound relative to the heme. If you need to, you can reset the molecules to their original orientation using the other button below the interactive window.
| Drag the structure with the mouse to rotate |
| Drag the structure with the mouse to rotate |
Notice the size of the active site in the CYP2E1 structure relative to that of the CYP3A4 structure. Although the size and shape of each cavity can change to accommodate different drugs, CYP2E1 cannot expand to the same degree that CYP3A4 can. In fact, CYP2E1 is known to only metabolize drugs that are quite small, such as ethanol, halothane, and aniline. On the opposite end of the spectrum, CYP3A4 can accommodate drugs that are quite large.
Focused QuestionsFocused Questions
- What is the reason that there is a cysteine residue in all isoforms of CYP450?
- What contributes to the fact that some drugs are metabolized at more than one location on the molecule?
- What contributes to the selectivity of one CYP isoform over another for metabolism of a given drug?
- What is "induced fit"?
- How is it possible for 2 co-administered drugs that are both substrates for a given CYP450 isoform to not apprently affect each other's metabolism?
- What interactions are important for binding of CYP1A2 to the isoflavone inhibitor? Are they lipophilic, ionic, or hydrogen bonds?
- What are the amino acid residues that surround the active site, and what kinds of interactions do these have with the drug?
- Why might water molecules be vital for binding of some drugs to some enzymes?
- What are 2 ways that a drug can inhibit CYP450 metabolism?
ReferencesReferences
- ↑ ISBN-13:978-1609133450
- ↑ Lynch T, Price A. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am Fam Physician. 2007 Aug 1;76(3):391-6. PMID:17708140
- ↑ ISBN-10:1609133455
- ↑ Sansen S, Yano JK, Reynald RL, Schoch GA, Griffin KJ, Stout CD, Johnson EF. Adaptations for the oxidation of polycyclic aromatic hydrocarbons exhibited by the structure of human P450 1A2. J Biol Chem. 2007 May 11;282(19):14348-55. Epub 2007 Feb 20. PMID:17311915 doi:10.1074/jbc.M611692200
- ↑ Yano JK, Wester MR, Schoch GA, Griffin KJ, Stout CD, Johnson EF. The structure of human microsomal cytochrome P450 3A4 determined by X-ray crystallography to 2.05-A resolution. J Biol Chem. 2004 Sep 10;279(37):38091-4. Epub 2004 Jul 16. PMID:15258162 doi:10.1074/jbc.C400293200
- ↑ Ekroos M, Sjogren T. Structural basis for ligand promiscuity in cytochrome P450 3A4. Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13682-7. Epub 2006 Sep 5. PMID:16954191
This page was originally created by users Kellan Passow, Arthur Cox, and Bob Hanson