Heme oxygenase
General InformationHeme Oxygenase (HO) is a member of the Hemoprotein family and catalyzes the Oxygen-dependent cleavage of the porphyrin ring of heme, using reducing equivalents like NADH to produce biliverdin, iron and CO [1]. HO consists of two main isoforms which are present in mammals, HO-1 and HO-2. The two isoforms are products of different genes, are different molecular sizes (32 kDa and 36 kDa respectively) and contain a different primary structure showing only 58% homology [2]. However studies have shown that the two isoforms share a region with 100% secondary structure homology which is believed to be the catalytic site of the protein[1]. The heme oxygenase isoforms are not free throughout the body but sequestered to certain tissues.
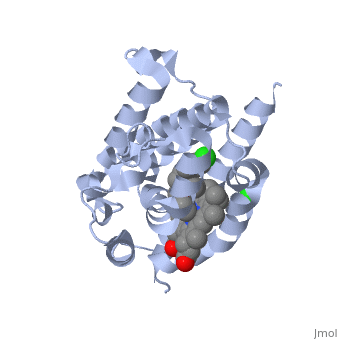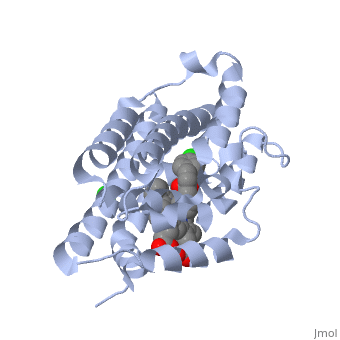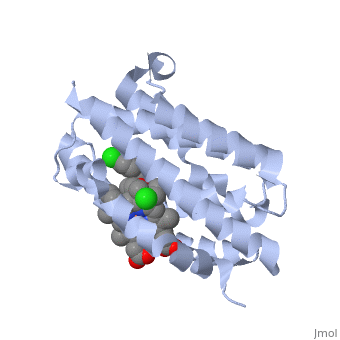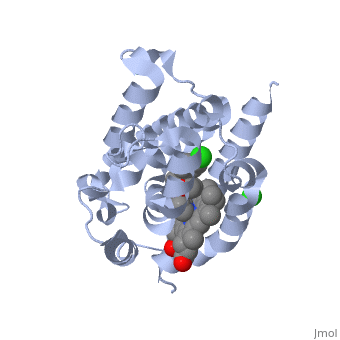
LigandHO non-covalently binds to a ligand known as a heme group more specifically Heme B. This Heme is the most abundant heme group commonly recognized for its role in oxygen transport and storage within mammalian tissues [4]. This group is contains a large heterocyclic ring known as a porphyrin ring with an iron atom in the center. The center iron atom serves as a source of electrons for the redox reaction to occur [4]. However this group is susceptible to damage from many stressors including physical shock and therefore needs to be broken down or recycled when theses stressors occur [1]. Structure
HO is a 233 residue protein with a secondary structure consisting of which interacts with a [1] at the optimum pH of 7.4; at 37 degrees C [5]. The heme is sandwiched between two helices termed the [6]. The proximal helix provides the His 25 heme ligand along with the various contact residues (Ala 28 and Glu 29), but also Thr 21 which contacts the heme through a water molecule [1]. On the distal side where the ligands binds (the catalytic site) there is a highly conserved sequence of Glycine residues () that provide a required flexibility for the reaction to occur [1]. This results in the backbone atoms of Gly 139 and Gly 143 to directly contact the heme. Inhibition of HO is provided by compounds such as imidazole-dioxolane which disrupt this flexibility, thereby forcing the HO protein to become rigid, stopping its function [7]. Of the four meso edges of the heme only one remains exposed () while the rest remain buried in the protein [2]. The exposed edge is the target of the HO reaction and requires the correct orientation for the hydroxylation reaction to occur[6]. This orientation is aided by the charges associated with the on the heme. The residues of are all near the propionates to anchor it via non convalent bonds so the a-meso carbon is in position for the hydroxylation reaction[8]. The propinates are located on the opposite side as the α-meso edge. The vinyl and methyl heme substituents do not appear to be important in orienting the heme due to the fact that they may be disordered about the y- axis which would change only the location of the methyl and vinyl groups while retaining the position of the propionates and the a-meso edge [8]. . Function Heme Oxygenase has two main functions, firstly it recycles iron supplies within the cell to maintain homeostasis and secondly it produces a product (biliverdin) that can be converted to a powerful antioxidant (bilirubin) which can aid in preventing oxidative cell damage [6]. The overall reaction consists of three sequential oxidation steps. In the first step the Oxygen bound to the heme iron is activated to become hydroperoxide. The production of α-hydroxyheme is accomplished by the electrophilic addition of its terminal oxygen to the α-meso carbon [6]. Then Heme Oxygenase converts α-hydroxyheme to verdoheme with the removal of the CO at the α-meso carbon (approximately 85% of CO produced under normal conditions is from this reaction)[6]. Lastly, the oxygen bridge of verdoheme is cleaved to produce biliverdin-iron chelate before the dissociation of the iron to biliverdin. (Figure 1) The electrons required for catalytic turnover of the enzyme are provided, in mammalian systems, by NADPH-cytochrome P450 reductase[6]. Studies suggest that the CO produced by the heme oxygenase reaction also functions to have anti-inflammatory, anti-proliferative and anti-apoptotic effects [7] to prevent cell damaage. A classic example of this reaction is a bruise. When tissue obtains a hard hit the erythrocytes release the heme creating a Red color. The heme then gets converted to biliverdin via heme oxygenase to produce a green color. Finally the conversion of biliverdin to bilirubin displays a yellow color. Therefore the reaction can be visibly observed [9]..
Medical Significance and Future Implications The lungs are a major target for various inflammatory, oxidative, carcinogenic and infectious pressures, which have the ability to result in a range of lung diseases like chronic obstructive lung diseases (COLD)[10]. The Induction of HO-1 is a crucial defense mechanism during these acute and chronic lung processes. The defense is obtained from the anti-oxidant, anti-inflammatory and anti-apoptotic properties of the products formed from the HO reaction [10]. (Figure 2) Therefore manipulation of the HO reaction and HO can have immense therapeutic potential against a range of lung diseases, if optimal levels of incorporation can be achieved. The products of the HO reaction are not just beneficial for lung diseases but also have a cytoprotective effect through the p38-MAPK pathway, and are a potential therapeutic treatment in cancer[1]. Recent studies have also shown that inhibition of the HO-1 reduces Kaposi sarcoma tumor growth [1]. Therefore future implications on regulating and manipulating this protein can have a massive impact on medical treatments. Additional ResourcesFor additional information, see: 3D structures of heme oxygenase
|
| ||||||||||
ReferencesReferences
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 Sugishima M, Higashimoto Y, Oishi T, Takahashi H, Sakamoto H, Noguchi M, Fukuyama K. X-ray crystallographic and biochemical characterization of the inhibitory action of an imidazole-dioxolane compound on heme oxygenase. Biochemistry. 2007 Feb 20;46(7):1860-7. Epub 2007 Jan 25. PMID:17253780 doi:10.1021/bi062264p
- ↑ 2.0 2.1 Lad L, Ortiz de Montellano PR, Poulos TL. Crystal structures of ferrous and ferrous-NO forms of verdoheme in a complex with human heme oxygenase-1: catalytic implications for heme cleavage. J Inorg Biochem. 2004 Nov;98(11):1686-95. PMID:15522396 doi:10.1016/j.jinorgbio.2004.07.004
- ↑ Otterbein LE, Soares MP, Yamashita K, Bach FH. Heme oxygenase-1: unleashing the protective properties of heme. Trends Immunol. 2003 Aug;24(8):449-55. PMID:12909459
- ↑ 4.0 4.1 Caughey WS, Smythe GA, O'Keeffe DH, Maskasky JE, Smith MI. Heme A of cytochrome c oxicase. Structure and properties: comparisons with hemes B, C, and S and derivatives. J Biol Chem. 1975 Oct 10;250(19):7602-22. PMID:170266
- ↑ Bonkovsky HL, Healey JF, Pohl J. Purification and characterization of heme oxygenase from chick liver. Comparison of the avian and mammalian enzymes. Eur J Biochem. 1990 Apr 20;189(1):155-66. PMID:2158889
- ↑ 6.0 6.1 6.2 6.3 6.4 6.5 Rahman MN, Vlahakis JZ, Szarek WA, Nakatsu K, Jia Z. X-ray Crystal Structure of Human Heme Oxygenase-1 in Complex with 1-(Adamantan-1-yl)-2-(1H-imidazol-1-yl)ethanone: A Common Binding Mode for Imidazole-Based Heme Oxygenase-1 Inhibitors. J Med Chem. 2008 Sep 18. PMID:18798608 doi:10.1021/jm800505m
- ↑ 7.0 7.1 Maines MD. Heme oxygenase: function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988 Jul;2(10):2557-68. PMID:3290025
- ↑ 8.0 8.1 Lee TS, Chau LY. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat Med. 2002 Mar;8(3):240-6. PMID:11875494 doi:10.1038/nm0302-240
- ↑ Evans JP, Niemevz F, Buldain G, de Montellano PO. Isoporphyrin intermediate in heme oxygenase catalysis. Oxidation of alpha-meso-phenylheme. J Biol Chem. 2008 Jul 11;283(28):19530-9. Epub 2008 May 16. PMID:18487208 doi:10.1074/jbc.M709685200
- ↑ 10.0 10.1 Raval CM, Lee PJ. Heme oxygenase-1 in lung disease. Curr Drug Targets. 2010 Dec;11(12):1532-40. PMID:20704548
This page originally authored by Barinder Chahal